
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 2 (2S): 19 – 22Special Issue–2 (Advances in Diagnosis and Control of Infectious Diseases of Animals)
Cloning and Sequence Analysis of Glycoprotein D Gene of Bovine Herpesvirus 1
Saurabh Majumder, Awadh Bihari Pandey, Muthannan Andavar Ramakrishnan*
-
Division of Virology, Indian Veterinary Research Institute, Muktheswar, Uttarakhand – 263 138
*Corresponding author:[email protected]
ARTICLE CITATION:
Majumder S, Pandey AB, Ramakrishnan MA, (2014). Cloning and sequence analysis of glycoprotein D gene of bovine herpesvirus 1. Adv. Anim. Vet. Sci. 2 (2S): 19 – 22.
Received: 2012–01–06, Revised: 2014–03–12, Accepted: 2014–03–13
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/journal.aavs/2014/2.2s.19.22
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Bovine herpesvirus 1 (BoHV1) infects mainly cattle and buffalo causing infectious bovine rhinotracheitis (IBR), infectious pustular vulvovaginitis (IPV) and infectious balanopostitis. It is also responsible for bovine respiratory disease complex or shipping fever along with other agents. It causes colossal economic losses due to productive and reproductive losses. Glycoprotein D is one of the most important surface glycoproteins of BoHV1. It induces strong neutralizing antibody response and it is the major protein involved in attachment, penetration and cell fusion. Glycoprotein D is a potential candidate for production of subunit vaccine. In this study amino (N) terminal 1041 bp of gD gene was amplified by using overlapping primers then the amplified fragments were cloned, sequenced and in–silico analysis was carried out. It was found that glycoprotein D of Indian isolate of BoHV1 is having 99.6% sequence identity with the reference sequence (Cooper strain, accession ID AJ004801) and less than 2% divergence with other BoHV1 strains. The deduced amino acid sequence analysis of gD showed a single amino acid change at position 313 which changed from glutamine (Cooper strain) to proline. BoHV1_IBR 216 II had divergence of 9.2%, 11.7% and 8.4% with BoHV–5, CvHV–1 and BuHV1, respectively. Phylogenetic analysis showed that this isolate under study forms same clad with other strains of BoHV1 around the world.
INTRODUCTION
Bovine herpesvirus 1 (BoHV1) belongs to genus Varicellovirus in the subfamily Alphaherpesvirinae, family Herpesviridae and order Herpesvirales. It infects mainly cattle (both domestic as well as wild) and buffalo but sheep and goats, swine, yak, mithun, mink and ferret are also affected by the virus (Qu and Li, 1988; Rajkhowa et al., 2004; Whetstone and Evermann, 1988). The virus causes latency and remains for undetected in the host for a long time in nervous ganglia. The disease is endemic in India. It causes huge economic losses mainly in feedlot cattle.
Glycoprotein D is a homolog of HSV1 gD (Tikoo et al., 1990), and one of the major glycoprotein present in the envelop of virus and plasma membrane of virus infected cell (van Drunen Littel–van den Hurk et al., 1984; Marshall et al., 1986). Its gene maps to the US region of the genome (US6) and encodes a polypeptide of 417 amino acids. It is essential for virus replication and is suggested to be involved in attachment, penetration (Chase and Letchworth, 1994; Fehler et al., 1992) and cell fusion (Tikoo et al., 1990). It induces strong neutralizing antibody response (Babiuk et al., 1987) and is recognized by both CD4+ and CD8+ T lymphocytes (Denis et al., 1993; Hutchings et al., 1990) making it a potential candidate for subunit vaccine and recombinant protein based ELISA.
Very few sequences are available for gD gene of Indian isolate of BoHV1 (Accession No. EU523745). For production of recombinant protein knowing the exact sequence of the gene is required. The present study describes the sequence analysis of amino (N) terminal 1041 bp of gD gene of BoHV1_IBR 216 II.
MATERIALS AND METHODS
Cells and Virus
Madin–Darby bovine kidney (MDBK) cells were grown in Dulbecco’s Modified Eagle Medium + GlutaMax–I (Invitrogen, USA) with 10% Newborn Calf Serum (Invitrogen, USA). The cells were incubated at 37 °C in the presence of 5% CO2. A BoHV1 isolate viz., (BoHV1_IBR 216 II) maintained in Division of Virology, IVRI, Mukteswar was used for the study.
Extraction of Viral DNA
The DNA was extracted using commercial kit as per the manufacture’s instruction (Promega, USA). In brief, MDBK cells infected with BoHV1 when showing 80% CPE were harvested. The infected cell layer was added with 200 μL of PBS and thrice freeze–thawed (30 min each at –80°C and 37°C). Then, 600 μL of nuclei lysis solution was added to the above lysate and mixed by pipeting. To remove RNA, 3 μL of RNase solution was added, mixed and incubated at 37°C for 30 min. About 200 μL of protein precipitation solution was added, vortexed and chilled on ice for 5 min. The suspension was centrifuged at 13000 xg for 4 min and the supernatant was transferred to a new tube. To precipitate the DNA, about 600 μL of isopropanol was added to the supernatant and centrifuged at 13000 xg for 1min. After decanting the supernatant, about 600 μL of 70% ethanol was added and centrifuging at 13000 xg for 1 min. After removing ethanol by aspiration and air drying, the DNA pellet was rehydrated with adding 100 μL of rehydration solution and incubating for 1 h at 65 °C.
Polymerase Chain Reaction
Oligonucleotides primers were designed based on sequence of the reference strain of BoHV1 (Genbank accession ID JX898220.1). Two pairs of overlapping primers were designed for amplification of glycoprotein D gene (Primer set 1 for amplification of fragment gD1, gD–F1–5’–CGC GTG TCT ATT GCG AGT TA –3’; gD–R1– 5’–GGC TCG TAG CCC TTC GACT –3 and primer set 2 for amplification of fragment gD2, gD–F2–5’–CGC GGA TCC GAT GAG CTT GCC TAC ACC–3’; gD–R2–5’–CCC AAG CTT TTA GTG CGT GAT GGC TTC GAG GCT–3’)’. The PCR reactions were carried out in a 50 µL of total volume of reaction mixture containing 25 μL of Maxima® Hot start Green PCR Master Mix (Fermentas, USA), 1 μL of forward primer, 1μL of reverse primer, 1 μL of template DNA and nuclease free water to 50 µL. The resulting mixture was subjected to a precise thermal profile in a programmable thermocycler as follows: One cycle at 95 °C for 5 minutes; 35 cycle at 95 °C for 1 minute, 55 °C for 1 minute, 72 °C for 1 minutes followed by one cycle at 72 °C for 10 minutes. Bands were visualized using ultraviolet transilluminator after running 6 µL of PCR products in 1% agarose gel. PCR amplicons of expected size was gel purified using gel purification kit (QIAquick® Gel Extraction Kit, Qiagen, USA) and used for downstream applications.
Cloning and Sequencing
The gel purified PCR amplicons was cloned using InsTA cloning Kit (Fermentas, USA). In brief, 1 μL of vector pTZ57R/T was mixed with 1 μL 10X Ligation buffer, 4 μL gel purified PCR product, 1 μL T4 DNA Ligase and nuclease free water was added to 10 μL. The mixture was vortexed, centrifuged for 3–5 sec and incubated for 2.5 h at 22 °C followed by for overnight at 4 °C. For transformation, 50 μL of competent DH5α from –80 °C was thawed on ice and Mixed with 5 μL of ligated PCR product and kept on ice for 30 min. The mixture was subjected to heat shock at 42 °C for 1 min, followed by 2 min on ice. 1 mL of LB broth was added to the tube and mixed by inversion, and incubated at 37 °C in a shaker incubator for 2 h. The cells were centrifuged at 5000 rpm for 2 min; the pellet was reconstituted in 50 μL of PBS and spreaded over LB agar plate containing ampicillin (50 μg/mL), X–gal (30 μg/mL) and IPTG (40 μg/mL). The plate was incubated at 37 °C for overnight at upright position. At least ten white colonies were selected and confirmed for having the desired construct by colony PCR using M13 primers. The confirmed bacterial containining positive inserts were commercially sequenced (SciGenome, Cochin, Kerala).
Editing of the raw sequence was done using BioEdit sequence alignment editor. References sequences of BoHV1 and other members of Alphaherpesvirinae were downloaded from GenBank. The Indian isolate in the current study and references sequences were aligned using the MEGA 5 software (Tamura et al., 2011). The phylogenetic tree was constructed using neighbor–joining algorithm with Kimura–2 parameters correction and 1000 bootstrap replications.
RESULT AND DISCUSSION
Bovine herpesvirus 1 is responsible for respiratory disease, abortion, decrease in milk production mainly in cattle and buffalo. It is an important cause of shipping fever and causes huge economic loss. Glycoprotein D is one of the important structural proteins of BoHV1. Molecular characterization of this gene will lay good foundation for further studies of this virus. This study glycoprotein D gene was characterized by nucleotide sequence analysis, deduced amino acid sequence analysis and phylogenetic analysis. This was done to compare Indian isolate IBR 216 II with other foreign isolates of BoHV1 and to identify potency of glycoprotein D for diagnosis, prophylaxis and epidemiological purpose.
On electrophoresis of PCR amplicon it formed expected size band of 939 bp (using first set primers, gD1) and 990 bp (using second set of primers, gD2) on agarose gel. After transformation ampicillin resistant colonies were subjected to colony PCR using universal M13 primers for confirmation of presence of the insert.
Figure 1: Result of colony PCR for confirmation of presence of PCR amplicon of gD gene using M13 universal primer
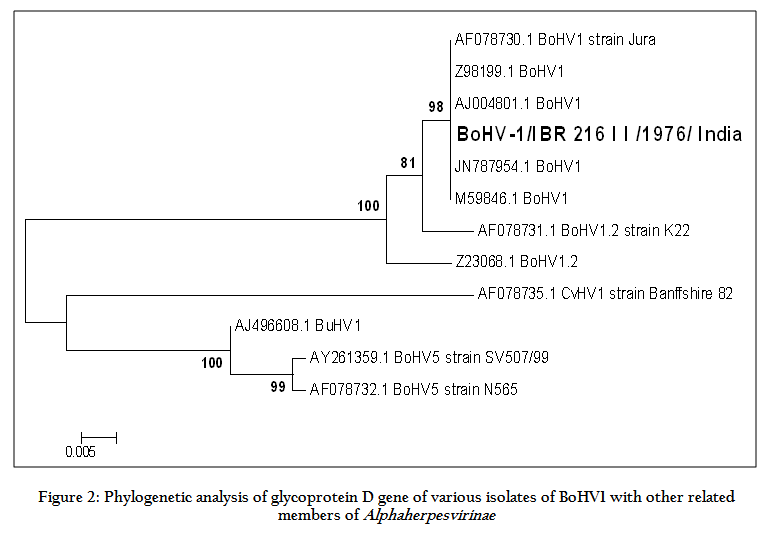
Figure 2: Phylogenetic analysis of glycoprotein D gene of various isolates of BoHV1 with other related members of Alphaherpesvirinae
We could sequence 1041 bp of the glycoprotein D using overlapping primers. The quality of DNA sequences was checked and overlapping fragments were assembled using the BioEdit. Assembled sequences with high quality were aligned using ClustalX. Homologies analyses were performed with the NCBI database and BLAST search for homologous sequences revealed sequence similarity between this ORF and the published gD gene of Alphaherpesviruses. Therefore the amplified gene is confirmed to be glycoprotein D. The N terminal portion of 1041 bp of gD gene was amplified, cloned and sequenced. Analysis of gD gene showed 99.6% sequence identity with the reference sequence and less than 2% divergence with other BoHV1 viruses. The BoHV1_IBR 216 II had divergence of 9.2%, 8.4% and 11.7% with BoHV5, BuHV1 and CvHV1, respectively. The GC content of the fragment is 68.6%. Phylogenetic analysis of gD gene was carried out for BoHV1_IBR 216 II with other BoHV1 and related groups of viruses. The analysis revealed that all BoHV1 viruses are clustered within single clade. There were 3 nucleotide changes found in this isolate with respect to the reference sequence those are at position 720 (guanine to adenine), 876 (cytosine to thymine) and 938 (adenine to cytosine). Whereas the deduced amino acid sequence analysis of gD showed a single amino acid change at position 313 which changed from glutamine (cooper strain) to proline. However, this change of one amino acid does not seem to affect the function of the protein. The N–glycosylation sites were found to be present at amino acid position 41 and 102 which is identical to that of the reference sequence.
CONCLUSION
This is the first genetic characterization of glycoprotein D gene (US6) of Indian isolate of BoHV1. Glycoprotein D gene of BoHV1 is highly conserved in all the isolates and it can be used as a target for designing of primers for diagnosis of IBR using PCR. As it is an immunodominant protein it can also serve as an ideal candidate of subunit vaccine production against BoHV1 infection.
ACKNOWLEDGEMENTS
The authors thank the Director, Indian veterinary research institute, Izatnagar, UP for providing the fund and the facilities to carry out the work. SM received fellowship from IVRI during the study.
REFERENCES
Babiuk LA, L'Italien J, van Drunen Littel–van den Hurk S, Zamb T, Lawman JP, Hughes G and Gifford GA (1987). Protection of cattle from bovine herpesvirus type I (BHV–1) infection by immunization with individual viral glycoproteins. Virology 159: 57–66.
http://dx.doi.org/10.1016/0042-6822(87)90347-3
Chase CC and Letchworth GJ 3rd (1994). Bovine herpesvirus 1 gIV–expressing cells resist virus penetration. J. Gen. Virol. 75 (Pt 1): 177–181.
http://dx.doi.org/10.1099/0022-1317-75-1-177
PMid:8113725
Denis M, Slaoui M and Keil G (1993). Identification of different target glycoproteins for bovine herpesvirus–1 specific cytotoxic T lymphocytes depending on the method of in vitro stimulation. Immunology 78: 7–13.
PMid:8382189 PMCid:PMC1421766
Van Drunen Littel–van den Hurk S, van den Hurk JV, Gilchrist JE, Misra V and Babiuk LA (1984). Interactions of monoclonal antibodies and bovine herpesvirus type 1 (BHV–1) glycoproteins: characterization of their biochemical and immunological properties. Virology 135: 466–479.
http://dx.doi.org/10.1016/0042-6822(84)90201-0
Fehler F, Herrmann JM, Saalmüller A, Mettenleiter TC and Keil GM (1992). Glycoprotein IV of bovine herpesvirus 1–expressing cell line complements and rescues a conditionally lethal viral mutant. J. Virol. 66: 831–839.
PMid:1309917 PMCid:PMC240783
Hutchings DL, van Drunen Littel–van den Hurk S and Babiuk LA (1990). Lymphocyte proliferative responses to separated bovine herpesvirus 1 proteins in immune cattle. J. Virol. 64: 5114–5122.
PMid:2168990 PMCid:PMC248003
Marshall RL, Rodriguez LL and Letchworth GJ 3rd (1986). Characterization of envelope proteins of infectious bovine rhinotracheitis virus (bovine herpesvirus 1) by biochemical and immunological methods. J. Virol. 57: 745–753.
PMid:3005625 PMCid:PMC252801
Qu XY and Li CH (1988). Studies on diagnosing infectious bovine rhinotracheitis in yak by ELISA. Chin J Vet Sci Tech 3: 5–8.
Rajkhowa S, Rajkhowa C, Rahman H and Bujarbaruah KM (2004). Seroprevalence of infectious bovine rhinotracheitis in mithun (Bos frontalis) in India. Rev Sci Tech 23: 821–829.
PMid:15861877
Tamura K, Peterson D, Peterson N, Stecher G, Nei M and Kumar S (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28: 2731–2739.
http://dx.doi.org/10.1093/molbev/msr121
PMid:21546353 PMCid:PMC3203626
Tikoo SK, Fitzpatrick DR, Babiuk LA and Zamb TJ (1990). Molecular cloning, sequencing, and expression of functional bovine herpesvirus 1 glycoprotein gIV in transfected bovine cells. J. Virol. 64: 5132–5142.
PMid:2168991 PMCid:PMC248005
Whetstone CA and Evermann JF (1988). Characterization of bovine herpesviruses isolated from six sheep and four goats by restriction endonuclease analysis and radioimmunoprecipitation. Am J Vet Res 49: 781–785.
PMid:2840840

