
Advances in Animal and Veterinary Sciences


Mini–Review Article
Advances in Animal and Veterinary Sciences. 1 (2S): 37 – 41Special Issue–2 (Clinical Veterinary Practice–T rends)
Endometrial Toll–like Receptors and Postpartum Uterine Infection in Bovine; a Minireview
Ajevar Ganesan3, Muthu Sankar1, Ramakrishnan Saravanan2, Goutam Kumar Das3, Harendra Kumar3, Arumugam Kumaresan4, Krishnaswamy Narayanan3*
- Division of Temperate Animal Husbandry, IVRI, Mukteswar campus,
- Immunology Section, IVRI, Izatnagar
- Division of Animal Reproduction, IVRI, IzatnagarD
- ivision of Livestock Production and Management, NDRI, Karnal, India
*Corresponding author:[email protected]
ARTICLE CITATION:
Ganesan A, Sankar M, Saravanan R, Das GK, Kumar H, Kumaresan A, Narayanan K (2013). Endometrial toll–like Receptors and postpartum uterine infection in bovine; a minireview. Adv. Anim. Vet. Sci. 2 (3):37 – 41.
Received: 2013–12–18, Revised: 2013–12–28, Accepted: 2013–12–31
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/241/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Postpartum uterine infection causes infertility and economic losses in the dairy industry. Since 2005, the host–pathogen interaction of uterine infection is investigated by understanding the innate uterine immunity through the interaction of endometrial Toll–like Receptors (TLRs) and the Pathogen associated molecular patterns (PAMPs) of the infectious agents. Transcriptional studies have confirmed the presence of most of the bubaline endometrial TLRs suggesting that the uterus is equipped to mount TLR mediated response following infection. However, unlike cattle, buffalo lacks the expression of endometrial TLR1. Association between the phases of estrous cycle with the specific TLR gene suggests that ovarian steroids regulate the expression which is up–regulated following uterine infections. Of all the TLRs, the interaction between Escherichia coli lipopolysaccharide (LPS) and TLR4 is best studied till date and shows activation of pro–inflammatory cytokines, preferential production of PGE2 and acute phase proteins in the bovine endometrium. Chronologically, intrauterine effects of LPS precede the discovery of TLRs. Emerging evidence indicates the presence of LPS in the follicular compartment in localized uterine infections; however, the vascular pathway carrying LPS of uterine origin to the ovary remains to be established. There is a scope for studying the other PAMPs of E coli with the cognate endometrial TLRs in the large ruminants. Similarly, no information on the interaction of PAMPS of recognized pathogens like Arcanobacterium pyogenes and Fusobacterium necrophorum with the corresponding endometrial TLRs is available. Expanding our understanding on the interaction of the common endometrial pathogens with the specific TLR in the endometrium is an obligatory pre–requisite for using specific TLR agonists in combating uterine infections.
The key determinants for excellent fertility in dairy cow herds is a healthy uterine environment, optimal estrus detection efficiency and ideal timing for breeding, since any afflictions of endometrium perturbs the normal reproductive functions leading to infertility or subfertility (Jabbour et al., 2009; Sheldon et al., 2009). The development of uterine disease is the equilibrium between the endometrial immune response and the virulence factors as well as the load of the causative microbe. Of the uterine diseases, postpartum (PP) metritis is by far, the most commonly encountered problem in the buffaloes (Raman and Bawa, 1977; Azawi, 2008). Apart from natural service, large scale implementation of artificial insemination coupled with unhygienic practice has resulted in an increased incidence of uterine infections in the buffaloes (Farca et al., 1997) thereby adversely affecting the fertility by delay in uterine involution, prolonged PP estrus, reduced conception rate and extended calving interval (Azawi, 2010). The occurrence and economic impact of clinical and subclinical endometritis (SE) in the PP cow has been the subject of many studies (LeBlanc et al., 2002; Kasimanickam et al., 2004; Gilbert et al., 2005; Hammon et al., 2006; Williams et al., 2007). Endometritis and associated inflammatory responses compromise the animal welfare, leading to infertility in the cows (Borsberry and Dobson, 1989; Sheldon and Dobson, 2004) and buffaloes (Usmani et al., 2001; Azawi, 2010). In India, it has been predicted that about 18–40% of cattle and buffaloes were culled mainly due to infertility (Sharma et al., 1993). The impact of abnormal calving on lactation yield and 305 days milk yield were hampered in buffaloes (Taraphder, 2002).
The innate immune response of the endometrium to pathogens involves an entire armory that includes complement system, antimicrobial peptides, immunoglobulins, acute phase proteins and pattern recognition receptors (PRRs) (Wira and Fahey, 2004). Germ–line–encoded PRRs recognize conserved pathogen–associated molecular patterns (PAMPs) synthesized by microorganisms but not by the host (Beutler, 2009; Takeuchi and Akira, 2010). The detection of PAMPs by PRRs in the endometrial cells triggers the production of cytokines and chemokines, which orchestrate an immune response to clear infection. The PRRs currently comprise four main classes of soluble and membrane–bound molecules viz., Toll–like receptors (TLRs), (RIG–I)–like receptors (RLRs), Nod–like receptors (NLRs) and C–type lectin receptors (CLRs) of which the most studied are the TLRs.
The TLRs belong to the subfamily Leucine–rich repeats (LRRs) that recognize conserved components of pathogens and are collectively called as pathogen associated molecular patterns (PAMPs) that initiate innate immune response. They belong to type I integral membrane glycoproteins and possess three major domains. The extracellular N–terminal domain consists of approximately 16–28 LRR and each LRR consists of 20–30 amino acids with the conserved motif (Kawai and Akira, 2010). Thus far, ten types of bovine TLRs have been identified and characterized (McGuire et al., 2005; Vahanan et al., 2008; Ganesan, 2013); each has its cognate ligands that activate specific intracellular signaling cascades resulting in upregulation of stimulus–specific cytokine(s) release to defend against invading pathogens.
(TLR2/TLRI or TLR2/TLR5)[H2] forms heterodimer to recognize an array of PAMPs predominantly from both Gram positive bacteria which includes lipopeptides, peptidoglycan, glycolipids, lipoteichoic acid (Kovacs–simon et al., 2011). The receptor dimerization is important, and tri–acetylated lipopeptides are usually bound by TLR2⁄TLR1, whereas di–acetylated lipopeptides are bound by TLR2⁄TLR6 (Jin et al., 2007). Recently, it has been shown that endometrial TLR2 and 6 are significantly upregulated in the buffalo with uterine infections (Ganesan, 2013). However, it remains to be elucidated whether the upregulation of those TLRs is due to the PAMPs of Gram–positive Archanobacterium pyogenes, the most common uterine pathogen.
Viral component of double stranded RNA (dsRNA) acts as a ligand for TLR3 and its synthetic analogue, polyinosinedeoxycytidylic acid I: C (Poly I: C) is widely used in in vitro studies. Uridine or guanosine–rich single–stranded RNAs from a wide range of viruses and synthetic imidazoquinoline–like molecules such as resiquimod are recognized by TLR7 and TLR8. Apart from LPS, use of other TLR agonists such as poly I: C and R848 in the treatment of endometritis has not yet been explored.
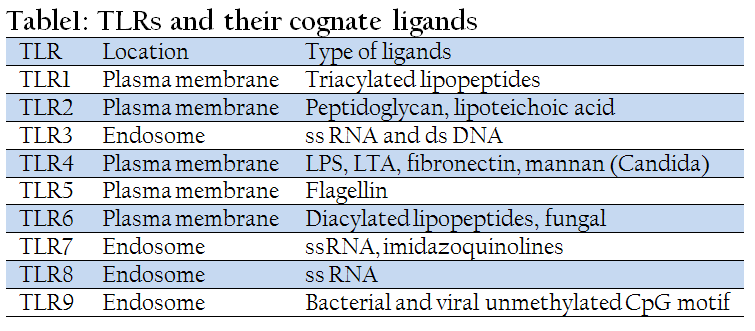
The LRR domains of TLRs are involved directly in the recognition of various components of microbes. The cognate ligands recognized by different TLRs are summarized in Table 1. Remarkably, despite the conservation among LRR domains, different TLRs can recognize several structurally unrelated ligands. The subcellular localization of different TLRs correlates to some extent with the molecular patterns of their ligands. TLR1, TLR2 and TLR4 are located on the cell surface and are recruited to phagosomes after activation by their respective ligands. By contrast, TLR3, TLR7 and TLR9, which are involved in the recognition of nucleic–acid–like structures, are not expressed on the cell surface. For instance, TLR9 has recently been shown to be expressed in the endoplasmic reticulum, and is recruited to endosomal/lysosomal compartments after stimulation with CpG motif (Akira et al., 2006).
Ligand recognition by TLRs leads to the recruitment of various TIR domain–containing adaptors such as myeloid differentiation primary–response protein 88 (MyD88), TIR–domain–containing adaptor protein (TIRAP), TIR–domain–containing adaptor protein inducing IFN–β (TRIF) and TRIF–related adaptor molecule (TRAM). The engagement of TLR1, TLR2, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9 and TLR11 with their respective ligands recruits MyD88. In addition to MyD88, TLR1, TLR2, TLR4 and TLR6 recruit TIRAP, which serves as a linker adaptor between the TIR domain of TLRs and MyD88. Binding of ligand with TLR3 and TLR4 recruits TRIF. Furthermore, TLR4 recruits TRIF through TRAM, which links between TIR domain of TLR4 and TRIF. This recruitment of adaptors triggers the cascade of signaling pathway and ultimately the activation of transcription factors such as nuclear factor–κB (NFκB) and interferon regulatory factor (IRF) (Kawai and Akira, 2010; Takeuchi and Akira, 2010).
From clinical point of view, the cellular components of E.coli like LPS and flagellum will be recognized by TLR4 and 5, respectively before it colonizes the endometrium. The cellular component of A.pyogenes such as triacylated lipoproteins and peptidoglycans are the potential ligands recognized by TLR2 ⁄TLR1 or TLR2⁄TLR6 (Kovacs–Simon, 2011). TLR9 recognizes the unmethylated CpG motifs of single–stranded DNA present in the genomes of many viruses and bacteria probably of IBR/IPVV of Herpes virus family and Brucella abortus as they are rich in CpG motif. Hence, the role of TLR agonists in clearing the uterine infection deserves investigation.
Contamination of bovine uterus is a consistent feature during parturition. Apart from bacterial load, virulence factors (VF) are the important pathogen–based determinants of infection. Bacterial overload, which is common following dystocia and retained fetal membranes, causes PP uterine infections (LeBlanc, 2008). Despite the fact that the culture of uterine samples yields a wide range of bacteria during the first 3 weeks PP, presence of Arcanobacterium pyogenes, Escherichia coli, Fusobacterium necrophorum and Provetella spp are commonly associated with clinical or subclinical endometritis (Griffin et al., 1974; Bonnett et al., 1991; Bicalho et al., 2010). E.coli isolated from bovine uterus within day 10 PP expressed a battery of VF, of which, fim H gene has been shown to increase the risk of endometritis (Silva et al., 2009; Bicalho et al., 2010). Analogous to enteropathogenic and uropathogenic E.coli, an endometrial pathogenic E. coli (EnPEC) has been characterized recently and was found to be more adherent and invasive to endometrial cells and stimulated production of chemokines and PGE2 in cell culture as well as mice model (Sheldon et al., 2010). Bicalho et al. (2010) reported that six E. coli VFs were associated with uterine infection: fimH, astA, cdt, kpsMII, ibeA, and hlyA. The VF gene fimH was the most prevalent in cows with metritis. Apart from E.coli, A. pyogenes has been emphasized in several studies as the main causative agent of endometrial damage and infertility (Dohmen et al., 2000). The VF that confers pathogenicity to A. pyogenes includes pyolysin (plo), a collagen–binding protein (CbpA), and a component of fimbriae (fimA). A hypothesis has been proposed that sequential infection of the PP uterus by the common bacteria determine the endometritis. For instance, E. coli infection during the first week of PP increases the susceptibility of the endometrium to subsequent infection with A. pyogenes (Olson et al., 1984; Williams et al., 2007), while A. pyogenes acts synergistically with F.necrophorum and Provetella sp to enhance the severity of uterine disease (Bonnett et al., 1991). Among the effects, E. coli releases bacterial cell–wall components like LPS (Williams et al., 2008); A. pyogenes produces the cholesterol–dependent cytotoxin, pyolysin (Bicalho et al., 2011) and a growth factor for F. necrophorum (Sheldon and Dobson 2004); F. necrophorum produces a leukotoxin; and Provetella sp produces a substance that inhibits phagocytosis (Sheldon and Dobson, 2004). Thus, apart from host factors, bacterial load and/or VFs of the common endometrial pathogens determine whether an infection of the PP uterus would result in recognized inflammatory conditions like clinical or subclinical endometritis. Beside the above bacteria, bovine herpes virus IV (BoHV–4) has been linked to cause uterine disease based on cell culture studies (Donofrio et al., 2007).
In cattle, the expression of TLR1 through 10 has been reported in the body of uterus, ipsilateral and contralateral horns (Davies et al., 2008). In buffalo, the expression profile of TLR2, 5, 7, 8, 9 and 10 in the normal uterus has been reported without delineating the influence of estrous cycle and discriminating endometrium from the myometrium (Vahanan et al.,2008). Barring Herath et al. (2006), others have studied the expression of TLRs only at transcriptional level. Immunohistochemical and western blot studies would throw much better information on the endometrial TLR expression. Further, in vitro and whole animal studies with agonist stimulation would indicate the functional competence of TLR proteins and unravel the immune response following ligand binding. Expression pattern of various endometrial TLRs is markedly influenced by the sex steroids owing to the functional dependency of endometrium across the estrous cycle. Hence, ovariectomized animal will be an appropriate model to study the endometrial TLRs expression as the hormonal milieu can be modulated by exogenous administration of estrogen and/or P4.
Transcripts of TLR4 and 5 showed highest expression during third trimester of pregnancy in cattle. Conversely, TLR 1/6 tend to show increased transcription during first trimester of gestation (Silva et al., 2012) suggesting that the uterus may be more responsive to pathogens during the period of pregnancy and subsequent parturition (Davies et al., 2008). An increased transcription of TLR 4 and 5 may be related to defense mechanism against such infections (King et al., 2003). In the inter–cotyledonary region, variable levels of TLR 1/6, TLR3, TLR4, TLR5, TLR7, TLR8, and TLR10 transcripts were observed (Martins et al., 2011). Oliveira et al. (2008) demonstrated that TLR9 has prominent role among the TLRs in attributing host resistance to B.abortus infection.
Most, if not all, the functional studies are confined to TLR4 as it recognizes the lipopolysaccharide component of E.coli. Administration of intravenous E. coli LPS disrupts neuroendocrine activity and results in interference with the estrous cycle. For instance, it is shown that LPS delayed ovulation, CL formation and rise in P4. In addition, upregulation of caspase–3 transcripts is also shown to compromise CL function and luteolysis (Herzog et al., 2012). Furthermore, CL diameter was smaller during the first PP cycle in cows with uterine pathogens suggesting an inimical effect of metritis on PP ovarian function (Williams et al, 2007). In our opinion, intravenous administration of LPS, at best, mimics the septic shock induced by toxic puerperal metritis that occur within day 10 PP and cannot be equated with PP metritis or subclinical endometritis as they are localized infections with little systemic involvement.
Cows with E. coli infection after parturition had more LPS and PGE in peripheral circulation. The association between infection and PGE was supported in vitro since LPS treatment of endometrial explants and cells preferentially stimulated PGE rather than PGF concentrations (Herath et al., 2009). It is well known that endometrial epithelial cells are the exquisite source of PGF as they harbor oxytocin receptors and stromal cells produce more PGE2 (Fortier et al., 2008). However, it is shown that endometrial epithelial cells accumulate more PGE than PGF in response to LPS. This switch in PG accumulation was associated with an increased level of phospholipase A2G6 (PLA2G6) protein in epithelial cells, rather than changes in the levels of PGES (PGE synthase) or PGFS (Herath et al., 2009). It would be interesting to know how LPS, or any infectious stimuli, cause a switch from PGF to PGE in the epithelial cells.
It has been demonstrated that LPS in the follicular fluid decreases the aromatase expression in the granulosa cells, increases the secretion of PGE and PGF, and transcripts of pro–inflammatory cytokine resulting in altered ovarian function (Sheldon and Bromfield, 2011; Borges et al., 2012; Jacca et al., 2013). However, uterine infection does not affect peripheral plasma FSH concentrations or the consequent emergence of a wave of growing follicles (Sheldon et al., 2002; Williams et al., 2007). The significance of localized uterine infection on the impairment of ovarian function is interesting as it confers LPS a ‘hormone–like role’; hence, it is necessary to delineate how the LPS reach follicular compartment or ovary from an infected uterus. In other words, the relative importance of utero–ovarian pathway and general circulation has to be unraveled in relation to uterine infections. In addition to LPS, E.coli possesses various VFs like fimbrae, cell wall proteins and adhesins which are putative ligands for various TLRs. For instance, presence of fim H gene in the uterus on PP day 3 is associated with risk of metritis (Bicalho et al., 2010). Hence, it is necessary to study the role of TLR5 vis–à–vis fimbriae (Davies et al., 2008). Practically, no data is available on the PAMPs of A. pyogenes, F necrophorum or Provetella sp with the corresponding bovine endometrial TLRs. Presence of LPS in the follicular fluid and its adverse effect on follicular and CL functions ushers us to investigate the presence and role of other bacterial components in the ovarian compartment. Since myometrial contractions play an important role in eliminating the exudate, the effects of TLR–PAMP interaction on inducing contraction associated proteins in the myometrium deserves attention. It is worth mentioning that before the advent of TLRs, intensive investigation with intrauterine LPS has been done as a treatment for endometritis in the cattle (Dhaliwal et al., 2001) and buffalo (Singh et al., 2000).
The uterus is exceptional among mucosal tissue on two accounts: first, ovarian steroid hormones have considerable effects on both afferent and efferent immune events (Wira and Rossoll, 1995). Thus, the outcome of an immune response and the elicitation of protective immunity to pathogens can be markedly influenced by the stage of the oestrous cycle at which priming or infection takes place. Second, uterus lacks highly organized secondary lymphoid nodules such as Peyer’s patch of the ileum (Robertson, 2000). In the cattle, innate immune response of endometrium under luteal phase is reduced and secretory activity of the endometrial glands provide congenial milieu for the establishment of pathogens. On the other hand, the rising levels of E2 in the follicular phase promote migration of immune cells into the uterine lumen, increasing uterine defenses (Liu et al., 2012).
The transcripts of TLR2/TLR6, TLR3, 4, 5, 6, 7 and 9 were found to be higher in the endometrium during the secretory phase than that of luteal phase of the menstrual cycle implying that P4 has a positive influence in the expression of TLRs (Nasu and Narahara, 2010).
CONCLUSION
Non–specific uterine infections during the postpartum period cause infertility and economic losses in the cattle and buffalo. The role of virulence factors in the establishment of infection by the common uterine microbes is emerging. Much of the expression studies on TLRs were done at mRNA level that needs to be corroborated at protein level by immunohistochemistry and Western blotting. Except LPS, other PAMPs of E.coli, A pyogenes, F necrophorum and Provetella have not been studied. Similarly, apart from TLR4, information on other TLRs is lacking. LPS released by the infected uterus reaches ovarian compartment and adversely affects the endocrine functions of follicle and CL. Regulation of endometrial TLRs by ovarian steroids need to be investigated ideally in ovariectomized model. Potential of different TLR agonists in combating uterine infection needs to be explored in the bovine.
REFERENCES
Akira, S, Uematsu, S. and Takeuchi, O. 2006. Pathogen recognition and innate immunity. Cell. 124:783 801.
http://dx.doi.org/10.1016/j.cell.2006.02.015
PMid:16497588
Azawi, O. I. 2010. Review: Uterine infection in buffalo cows. Buffalo Bulletin. 29: 154–171.
Azawi, O.I. 2008. Postpartum uterine infection in cattle. – review. Animal Reproduction Science.105: 187–208.
http://dx.doi.org/10.1016/j.anireprosci.2008.01.010
PMid:18280065
Beutler, B, A. 2009. TLRs and innate immunity. Blood 113:1399–407.
http://dx.doi.org/10.1182/blood-2008-07-019307
PMid:18757776 PMCid:PMC2644070
Bicalho, M.L., Machado, V.S., Oikonomou, G., Gilbert, R.O., Bicalho, R.C. 2012. Association between virulence factors of Escherichia coli Fusobacterium Necrophorum, and Arcanobacterium Pyogenes and uterine diseases of dairy cows. Veterianry Microbiology. 157: 125–13.
http://dx.doi.org/10.1016/j.vetmic.2011.11.034
PMid:22186615
Bicalho, R.C., Machado, V.S., Bicalho, M.L., Gilbert, R.O., Teixeira, A.G., Caixeta, L.S., Pereira, R.V. 2010. Molecular and epidemiological characterization of bovine intrauterine Escherichia coli. Journal of Dairy Science. 5818–5830.
http://dx.doi.org/10.3168/jds.2010-3550
PMid:21094754
Bondurant, R.H., 1999. Inflammation in the bovine female reproductive tract. Journal of Animal Science. 77: 101–110.
PMid:15526785
Bonnett, B. N., Martin, S. W., Gannon, V. P. G., Miller, R. B and Etherington, W. G. 1991. Endometrial biopsy in Holstein– Frisian dairy cows. III. Bacteriological analysis and correlation with histopathological findings. Canadian Journal of Veterinary Research, 55: 168–173.
PMid:1884297 PMCid:PMC1263438
Borges, A. M., Healey, G. D and Sheldon, I. M. 2012. Explants of intact endometrium to model bovine innate immunity and inflammation ex vivo. American Journal of Reproductive Immunology 67: 526–539.
http://dx.doi.org/10.1111/j.1600-0897.2012.01106.x
PMid:22324889
Borsberry, S and Dobson, H. 1989. Periparturent diseases and their effect on reproductive performance in five dairy herds. Veterinary Record. 124: 217–219.
http://dx.doi.org/10.1136/vr.124.9.217
PMid:2929110
Davies, D., Meade, K. G., Herath, S., Eckersall, P. D., Gonzalez, D., White, J. O., Conlan, R. S., O'Farrelly, C. and Sheldon, I. M. 2008. Toll–like receptor and antimicrobial peptide expression in the bovine endometrium. Reproductive Biology and Endocrinology. 6: 1–12.
http://dx.doi.org/10.1186/1477-7827-6-53
PMid:19017375 PMCid:PMC2627908
Dhaliwal, G. S., Murray, R. D. and Woldehiwet, Z. 2001. Some aspects of immunology of the bovine uterus related to treatments for endometritis. Animal Reproduction Science 67: 135–152.
http://dx.doi.org/10.1016/S0378-4320(01)00124-5
Dohmen, M. J. W., Joop, K., Strurk, A., Bols, P. E. J. and Lohuis, J. A. C. M. 2000. Relationship between intra–uterine bacterial contamination endotoxin levels and the development of endometritis in postpartum cows with Dystocia or retained placenta. Theriogenology. 54: 1019–1032.
http://dx.doi.org/10.1016/S0093-691X(00)00410-6
Donofrio, G., Herath, S., Sartori, C., Cavirani, S., Flammini, C. and Sheldon, I.M. 2007. Bovine Herpesvirus 4 (BoHV–4) is tropic for Bovine endometrial cells and modulates endocrine function. Journal of Reproduction and Fertility. 134: 183–197.
http://dx.doi.org/10.1530/REP-07-0065
PMid:17641100 PMCid:PMC2740819
Farca, A. M., Nebbia, P., Robino, P. and Re, G. 1997. Effects of the combination antibiotic –EDTA–TRIS in the treatment of chronic bovine endometritis caused by the anti–microbial–resistant bacteria. Pharmacology Research. 36: 1–6.
http://dx.doi.org/10.1006/phrs.1997.0199
PMid:9368912
Forteir,M.A., Narayanan,K., Danyod,G., Kovalik,S.B., and Chapdelaine,J.A. 2008. A postgenomic integrated view of prostaglandins in reproduction:implications for other body systems. Journal of Physiology and Pharmacology. 59:65–89
Gilbert, R. O., Shin, S. T., Guard, C. L., Erb, H. N. and Frajblat, M. 2005. Prevalence of endometritis and its effect on reproductive performance of dairy cows. Theriogenology. 64: 1879–1888.
http://dx.doi.org/10.1016/j.theriogenology.2005.04.022
PMid:15961149
Griffin, J. F. T., Hartigan, P. J. and Nunn, W. R. 1974. Non–specific uterine infection and bovine fertility I: infection patterns and endometritis during the first seven weeks postpartum. Theriogenology. 1: 91–106.
http://dx.doi.org/10.1016/0093-691X(74)90053-3
http://dx.doi.org/10.1016/0093-691X(74)90052-1
Hammon, D. S., Evjen, I. M., Dhiman, T. R., Goff, J. P. and Walters, J. L.2006. Neutrophil function and energy status in Holstein cow with uterine health disorders. Veterinary Immunology Immunopathology. 113: 21–9.
http://dx.doi.org/10.1016/j.vetimm.2006.03.022
PMid:16740320
Herath, S., Fischer, D. P., Erin, D. W., Williams, J., Lilly, S. T., Dobson, H., Bryant, C. E. and Sheldon, I.M. 2006. Expression and function of toll–like receptor 4 in the endometrial cells of the uterus. Endocrinology. 147: 562–570.
http://dx.doi.org/10.1210/en.2005-1113
PMid:16223858 PMCid:PMC2738982
Herath, S., Lilly, S.T., Fischer, D.P., Williams, E.J., Dobson, H., Bryant, C.E. and Sheldon,I.M. 2009.Bacterial lipopolysaccharide induces an endocrine switch from prostaglandin F2alpha to prostaglandin E2 in bovine endometrium. Endocrinology. 150: 1912–1920.
http://dx.doi.org/10.1210/en.2008-1379
PMid:19056817 PMCid:PMC2706387
Herzog, K., Struve, K., Kastelic, J, P., Piechotta, Y., Ulbruich, S, E., Pfarrer, C., Shirasuna, K., Shimizu, T., Miyamoto, A. and Bollwein, H. 2012. Escherichia coli lipopolysaccharide administration transiently suppresses luteal structure and function in diestrous cows. Journal of Reproduction and Fertility. 144: 467–476.
http://dx.doi.org/10.1530/REP-12-0138
PMid:22829687
Hirata, T., Osuga, Y., Hirota, Y., Koga, K., Yoshino, O., Harada, M., Morimoto, C., Yano, T., Nishii, O., Tsutsumi, O. and Taketani, Y. 2005. Evidence for the presence of toll– like receptor 4 system in the human endometrium. Journal of Clinical Endocrinology and Metabolism. 90: 548–556.
http://dx.doi.org/10.1210/jc.2004-0241
PMid:15509642
Jabbour, H. N., Sales, K. J., Catalano, R. D., Norman, J. E. 2009. Inflammatory pathways in female reproductive health and disease. Reproduction 138: 903–919.
http://dx.doi.org/10.1530/REP-09-0247
PMid:19793840
Jacaa, S., Franceschi, V., Colagiorgi, A., Sheldon, M., and Donofrio, G. 2013. Bovine endometrial stromal cells support Tumor necrosis factor alpha–induced bovine herpes virus type 4 enhanced replication. DOI: 10.1095/1 Biology of Reproduction. 112.106740.
Jin, M. S., S. E. Kim, J. Y. Heo, M. E. Lee, H. M. Kim, S. G. Paik, H. Lee, and J. O. Lee. 2007. Crystal structure of the TLR1–TLR2 heterodimer induced by binding of a tri–acylated lipopeptide. Cell 130:1071–1082.
http://dx.doi.org/10.1016/j.cell.2007.09.008
PMid:17889651
Kasimanickam, R., Duffield, T. E., Foster, R. A., Gartley, C. L., Leslie, K. E., Walton, J. S., Johnson, W. H. 2004. Endometrial cytology and ultrasonography for the detection of sub clinical endometritis in postpartum dairy cows. Theriogenology. 62: 9–23.
http://dx.doi.org/10.1016/j.theriogenology.2003.03.001
PMid:15159097
Kawai, T. and Akira, S. 2010. The role of pattern–recognition receptors in innate immunity: update on Toll–like receptors. Nature Immunology.11: 373–84.
http://dx.doi.org/10.1038/ni.1863
PMid:20404851
King, A. E., Critchley, H. O. and Kelly, R. W. 2003. Innate immune defences in the human endometrium. Reprodcutive Biology and Endocrinology. 1: 116.
http://dx.doi.org/10.1186/1477-7827-1-116
PMid:14641912 PMCid:PMC305332
Kovacs–simon, A., Titball,W.R., and Michell,L.S. 2011. Lipoproteins of bacterial pathogens. Infection Immunity 79:548–561
http://dx.doi.org/10.1128/IAI.00682-10
PMid:20974828 PMCid:PMC3028857
LeBlanc, S. J. D., Duffield, T. F., Leslie, K. E., Bateman, K. G., Keefe, G. P., Walton, J. S. and Johnson, W. H. 2002. The effect of treatment of clinical endometritis on reproductive performance in dairy cows. Journal of Dairy Science. 85: 2237–2249.
http://dx.doi.org/10.3168/jds.S0022-0302(02)74303-8
http://dx.doi.org/10.3168/jds.S0022-0302(02)74302-6
LeBlanc, S. 2008.Postpartum uterine disease and dairy herd reproductive performance – a review. The Veterinary Journal. 176: 102–114.
http://dx.doi.org/10.1016/j.tvjl.2007.12.019
PMid:18328749
Lewis, G. S. 1997. Health problems of the postpartum cow, uterine health and disorders. Journal of Dairy Science. 80: 984–994.
http://dx.doi.org/10.3168/jds.S0022-0302(97)76024-7
Liu.S., Zuo, Z.,Yang,Z., Liu, B., and Zhang,N. 2012. Roles of toll–like receptor 4 and immunomodulatory hormones in bovine endometritis. African Journal of Pharmacy and Pharmacology. 6(15) 1136 – 1141
Martins, T.M., Paixão, T.A., Costa, E.A., Pires, A.C., Santos, R.L., Borges, Á.M. 2011. Postpartum toll–like receptors and beta–defensin 5 mRNA levels in the endometrium of Holstein cows. Veterinary immunology and immunopathology. 139: 277–281.
http://dx.doi.org/10.1016/j.vetimm.2010.09.012
PMid:21067816
McGuire, K., Jones, M, F, D., Williams, J. L., Glass, E. J. and Jann, O. 2005. Radiation hybrid mapping of all 10 characterized bovine Toll–like receptors. Animal Genetics. 37: 47–50.
http://dx.doi.org/10.1111/j.1365-2052.2005.01364.x
PMid:16441295
Nasu, K. and Narahara, H. 2010. Pattern recognition via the Toll like receptor system in the human female genital tract. Mediators of Inflammation Doi:10.1155/976024.
Oliveira, S, C., de Oliveira, F, S., Macedo, G, C., de Almeida, L, A., Carvalho, N, B. 2008. The role of innate immune receptors in the control of Brucella abortus infection: toll–like receptors and beyond. Microbial Infection. 10(9): 1005–9.
http://dx.doi.org/10.1016/j.micinf.2008.07.005
PMid:18664388
Olson, J. D., Ball, L., Mortimer, R. G., Farin, P, W., Adney, W. S. and Huffman, E. M. 1984. Aspects of bacteriology and endocrinology of cows with pyometra and retained fetal membranes. American Journal of Veterinary Research. 45: 2251–2255.
PMid:6524717
Raju, K. G. S., Naidu, V. K. and Rao, V. K. 2007. Incidence of reproductive disorders in buffalo under field conditions of Andhra Pradesh. Indian Journal of Animal Reproduction. 28: 46–48.
Raman, S. R. P. and Bawa, S. J. S.1977.Incidence of pre and postpartum reproductive disorders in buffaloes. Haryana Veterinarian,16:99–101.
Robertson, S. 2000. Control of the immunological environment of the uterus. Journal of Reproduction and Fertility. 5: 164–174.
Sharma, S. S., Gupta, A. K., Bishnoi, B. L., Pareek, P. K, and Rawat, M. 1993. Antibiotic sensitivity patterns of microorganisms causing endometritis in cattle. Indian Journal of Animal Reproduction. 14: 116–117.
Sheldon, I. M. and Bromfield, J. J. 2011. Innate immunity in the human endometrium and ovary. American Journal of Reproductive Immunology. 66(1): 63–71.
http://dx.doi.org/10.1111/j.1600-0897.2011.01034.x
PMid:21726339
Sheldon, I. M., Price, S. B., Cronin, J., Gilbert, R. O. and Gadsby, J. E. 2009. Mechanism of infertility associated with clinical and subclinical endometrtitis in high producing dairy cattle. Reproduction in Domestic Animal. 44: 1–9.
http://dx.doi.org/10.1111/j.1439-0531.2009.01465.x
PMid:19660075
Sheldon, I. M., Noakes, D. E., Rycroft, A. N., Pfeiffer, D. U., Dobson, H. 2002. Influence of uterine bacterial contamination after parturition on ovarian dominant follicle selection and follicle growth and function in cattle. Journal of Reproduction and Fertility. 123: 837–845.
http://dx.doi.org/10.1530/rep.0.1230837
Sheldon, I. M., Rycroft, A. N., Dogan, B, Craven, M., Bromfield, J. J., Chandler, A., Roberts, M. H., Price, S, B., Gilbert, R. O. and Simpson, K. W. 2010. Specific strains of Escherichia coli are pathogenic for the endometrium of cattle and cause pelvic inflammatory disease in cattle and mice. PLoS One. 5: e9192.
http://dx.doi.org/10.1371/journal.pone.0009192
PMid:20169203 PMCid:PMC2820550
Sheldon,I.M. and Dobson, H. 2004. Postpartum uterine health in cattle. Animal Reproduction Science 82–83: 295–306.
http://dx.doi.org/10.1016/j.anireprosci.2004.04.006
PMid:15271461
Silva, C. P. A., Costaa, A, E. and Macêdoa, A, A., Martinsa, M. T. D., Borges, M. A, Paixãoba, T., Santosa, R. 2012.Transcription of pattern recognition receptors and abortive agents induced chemokines in the bovine pregnant uterus. Veterinary Immunology and Immunopathology 145 : 248–256.
http://dx.doi.org/10.1016/j.vetimm.2011.11.007
PMid:22153993
Silva, E., Leitao, S., Tenreiro, T., Pomba, C., Nunes, T., Lopes da Costa, L., Mateus, L. 2009. Genomic and phenotypic characterization of Escherichia coli isolates recovered from the uterus of puerperal dairy cows. Journal of Dairy Science 92: 6000–6010.
http://dx.doi.org/10.3168/jds.2009-2358
PMid:19923603
Singh, J., Sidhu, S. S., Dhaliwal, G. S., Pangaonkar, G. R., Nanda, A. S., Grewal, A. S. 2000. Effectiveness of lipopolysaccharide as an intrauterine immunomodulator in curing bacterial endometritis in repeat breeding cross–bred cows. Animal Reproduction science. 59: 159–166.
http://dx.doi.org/10.1016/S0378-4320(00)00144-5
Takeuchi, O. and Akira, S. 2010. Pattern recognition receptors and inflammation. Cell. 140: 805–820.
http://dx.doi.org/10.1016/j.cell.2010.01.022
PMid:20303872
Taraphder, S. 2002. Genetic and economic evaluation of Murrah buffaloes for lactation disorders and disposal pattern. Ph.D. thesis submitted to the National Dairy Research Institute, Karnal.
Usmani, R. H., Ahmad, N., Shafiq, P. and Mirza, M. A. 2001. Effect of sub clinical uterine infection on cervical and uterine involution, estrous activity and fertility in postpartum buffaloes. Theriogenology. 55: 563–571.
http://dx.doi.org/10.1016/S0093-691X(01)00426-5
Vahanan, M. B., Raj, G. D., Pawar, R. M. C., Gopinath, V. P., Raja, A., Thangavelu, A. 2008. Expression profile of toll like receptor in a range of water buffalo tissues (bubalus bubalis). Veterinary Immunology and Immunopathology. 126: 149–155.
http://dx.doi.org/10.1016/j.vetimm.2008.05.027
PMid:18619678
Williams, E. J., Fischer, D. P., Noakes, D. E., England, G. E., Rycroft, A., Dobson, H. and Sheldon, I. M. 2007. Uterine infection perturbs ovarian function in the postpartum dairy cow. Theriogenology. 68: 549–559.
http://dx.doi.org/10.1016/j.theriogenology.2007.04.056
PMid:17574659 PMCid:PMC2702080
Williams, E. J., Sibley, K., Miller, N. A., Fishwick, J., Herath, S., Deborah, England. C. W., Dobson, H., Sheldon. I. M. 2008. The effects of Escherichia coli lipopolysaccharide and TNF alpha on ovarian function, American Journal of Reproductive Immunology. 60(5): 462–473.
http://dx.doi.org/10.1111/j.1600-0897.2008.00645.x
PMid:19238751 PMCid:PMC2702079
Wira, C. R. and Rossoll, R. M. 1995.Antigen–presenting cells in the female reproductive tract: influence of sex hormones on antigen presentation in the vagina. Immunology. 84: 505–508.
PMid:7790022 PMCid:PMC1415167
Wira, C. R. and Fahey, J. V. 2004 .The innate immune system: gatekeeper to the female reproductive tract. Immunology. 111: 13–15.
http://dx.doi.org/10.1111/j.1365-2567.2004.01796.x
PMid:14678193 PMCid:PMC1782397

