
Influence of Protein-Energy Balance on Growth, Production, and Reproduction in Pesisir Heifers



Research Article
Influence of Protein-Energy Balance on Growth, Production, and Reproduction in Pesisir Heifers
Roni Pazla1, Mardiati Zain1*, Fauzia Agustin1, Yetti Marlida1, Zaituni Udin2, Jaswandi2, Masrizal2, Hendri2, Windu Negara3, Totti Tjiptosumirat3, Ezi Masdia Putri3, Multiviza Muslim4
1Department of Nutrition and Feed Technology, Faculty of Animal Science, Universitas Andalas, Padang, 25163, West Sumatera, Indonesia; 2Department of Livestock Production Technology, Faculty of Animal Science, Universitas Andalas, Padang, 25163, West Sumatera, Indonesia; 3Research Center for Animal Husbandry, Research Organization for Agriculture and Food, National Research and Innovation Agency (BRIN), Jl. Raya Jakarta-Bogor Cibinong 16915, Indonesia; 4Center for Breeding Superior Livestock and Forage for Animal Feed Padang Mengatas, Jl. Raya Payakumbuh-Lintau, KM.9 Pekan Sabtu, Payakumbuh 26201, Indonesia.
Abstract | The protein-energy ratio in Pesisir cattle diets plays a role in determining their productivity. We aimed to establish the most effective combination of crude protein (CP) and total digestible nutrients (TDN) in a ration to enhance the productivity of Pesisir heifers. We evaluated consumption, nutrient digestibility, production performance, and the percentage of first estrous in sixteen heifers at 12-15 m and averaging 100.09 kg in a randomized block design with a 2 x 2 x 4 factorial pattern, where factor A represented crude protein, and factor B represented the TDN level of the ration. The treatment combinations were A1B1 (CP 10% and TDN 60%), A1B2 (CP 10% and TDN 65%), A2B1 (CP 12% and TDN 60%), and A2B2 (CP 12% and TDN 65%). The results revealed a significant interaction between factors concerning consumption and nutrient digestibility. However, there was no significant interaction for production performance. Notably, the highest percentage of first estrous was found in heifers that received A1B2. A diet with 10% CP and 65% TDN could increase consumption, feed digestibility, production performance, and the estrous number of Pesisir heifers.
Keywords | Crude protein, Estrous, Performance, Pesisir Heifers, TDN
Received | November 02, 2023; Accepted | December 15, 2023; Published | February 12, 2024
*Correspondence | Mardiati Zain, Department of Nutrition and Feed Technology, Faculty of Animal Science, Universitas Andalas, Padang, 25163, West Sumatera, Indonesia; Email: mardiati@ansci.unand.ac.id
Citation | Pazla R, Zain M, Agustin F, Marlida Y, Udin Z, Jaswandi, Masrizal, Hendri, Negara W, Tjiptosumirat T, Putri EM, Muslim M (2024). Influence of protein-energy balance on growth, production, and reproduction in Pesisir heifers. Adv. Anim. Vet. Sci., 12(3): 546-558.
DOI | https://dx.doi.org/10.17582/journal.aavs/2024/12.3.546.558
ISSN (Online) | 2307-8316
Copyright: 2024 by the authors. Licensee ResearchersLinks Ltd, England, UK.
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
INTRODUCTION
Cows are ruminant livestock that supply human protein needs and play a significant role in meeting both national and international demands for animal protein (Basyar, 2021). Pighin et al. (2016) stated that beef remains a popular choice due to its complete nutritional profile, including protein and fat, and its appealing taste. Pesisir cattle, a local Indonesian bread, exhibit high adaptability to low-quality feed and resistance to various diseases and parasites (Hartatik et al., 2018). They demonstrate superior heat tolerance in hot environments compared to the simmental and ongole crossbreeds (Yetmaneli et al., 2023). This adaptability in tropical regions is influenced by the presence of large sweat glands and their density, providing them with advantages in thermoregulation.
Recently, the development of Pesisir cattle has encountered challenges resulting in a decline in production. This decline is evident in both the population and productivity, attributed to limited environmental carrying capacity and competition from imported cattle. In 2020, the Pesisir cattle population recorded 85,031 individuals. This figure marked a significant decrease compared to the population in 2008, which reached 89,995 heads, with a decline rate of 5.84% (BPS, 2020). Pesisir cattle have a relatively high carcass percentage (50.6%) compared to buffalo, Ongole crossbreed, Madurese, and Ongole cattle. They rank second to Balinese cattle, which have a percentage of 56.9% (Hendri, 2013). One contributing factor to the suboptimal productivity of Pesisir cattle is the provision of feed with low nutritional content and inadequate community maintenance management. There is an urgent need for initiatives to boost the Pesisir cattle population through feed engineering, with a particular emphasis on Pesisir heifers, as they are prospective parent cattle.
The nutrients that livestock need are protein and energy (Pazla et al., 2023a). Protein plays a role in facilitating the growth or formation of body cells, particularly in calves and young cattle. Livestock experiencing nutritional deficiencies will encounter reduced reproductive efficiency and reproductive disorders, affecting both females and males. A deficiency in protein within the feed can lead to reproductive disorders in livestock, such as weak estrous, embryo death, and premature birth. The balance between protein and TDN in the ration significantly influences the efficiency of nutrient utilization, ultimately impacting livestock productivity (Pazla et al., 2021a). The balance of protein and TDN plays a crucial role in formulating cattle rations, if this balance is not maintained, it can result in an excess or deficiency of energy and protein intake in the animals. Nugroho et al. (2013) stated that there is a substantial need for a balance of protein and TDN, especially in young ruminants during their growth phase. We aimed to identify the optimal protein and TDN balance for Pesisir heifers concerning feed consumption, nutrient digestibility, body weight gain, production performance, and the percentage of first estrous.
MATERIALS AND METHODS
Experimental location
The trial was carried out at the Center for Breeding Superior Livestock and Forage for Animal Feed in Padang Mengatas, located at Jl. Raya Payakumbuh-Lintau, KM.9, Pekan Sabtu, Payakumbuh 26201, Indonesia, spanning from May to September 2023. The altitude of this area is 790-1014 m above sea level. The region experiences an average annual rainfall of 1800 mm, with temperatures between 18 and 28°C, and a relative humidity of around 70%. Watering source is from around the cage and ad bilitum availability. The cattle was feed with concentrate in the morning and forage was given twice in a day. The cage has good drainage and ventilation system. Lighting was provided in the night. The cage was cleaned everyday and also the cage and the person were sprayed with disinfectant. Anthelmintic procedure was given before experiment.
Research materials and methods
This research used a 2 x 2 factorial randomized block design method with four replications. Factor A represented protein content at 10 and 12%, while factor B represented TDN content at 60 and 65%. We used sixteen Pesisir heifers aged 12-15 months with an average body weight of 100.09± 14.85 kg. the animals were divided into four groups of four Pesisir heifers as follows: GI at 73.5-84.5 kg (20.03%), GII at 86-94 kg (16.07%), GIII at 106.5- 112.5 kg (15.37%) and GIV at 115.5-119.5 (15.76%). The feed treatments were A1B1 (10% CP and 60% TDN), A1B2 (12% CP and 60% TDN), A2B1 (10% CP and 65% TDN), and A2B2 (12% CP and TDN 65%).
This research comprised four periods. The 1st is the Adaptation Period, which spanned 10 days to acquaint Pesisir heifers with new environmental conditions and enable them to adapt to the forthcoming treatment rations. The 2nd is the observation period (Preliminary), a 15-day designed to eliminate any influence from previous rations. The 3rd is the collection period of 5 days aimed to assess the digestibility of the ration via a collection method. During this period, daily measurements were taken for ration consumption, feces, and urine excretion. Approximately 10% of the daily feces and urine were collected and stored in designated containers. The feces were air-dried using Memmert Universal Oven Model UN55 from Germany, followed by an additional oven-drying in an oven at 60°C for 48 hours. Nutritional contents in food and feces samples were analyzed using proximate analysis based on the Association of Official Agricultural Chemists (2016), and fiber fractions were determined using the method developed by Van Soest et al. (1991). The 4th is the growth period for two months post-collection period, which involves measuring the initial and final body weights of the Pesisir heifers. Estrous was observed during the study by monitoring signs of estrous, where vaginal discharge indicated a normal cycle. The percentage of estrus cows was calculated as follows:
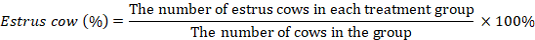
The experimental diet was composed of a 60:40 ratio of forage to concentrate. Elephant grass was used as the forage, while the concentrate was comprised of bran, corn, palm kernel meal, and a blend of minerals for cattle. The specific composition of the feed ingredients in the diet can be found in Table 1, and the chemical composition of the diet is detailed in Table 2.
Table 1: Ration composition of treatment ration.
|
Composition (%) |
A1B1 |
A1B2 |
A2B1 |
A2B2 |
|
Corn |
7 |
25 |
2 |
17 |
|
Palm Kernel cake |
6 |
2 |
15 |
14 |
|
Mineral |
1 |
1 |
1 |
1 |
|
NaCl (table salt) |
0.5 |
0.5 |
0.5 |
0.5 |
|
Rice bran |
25.5 |
11.5 |
21.5 |
7.5 |
|
Elephant grass |
60 |
60 |
60 |
60 |
A1B1 (CP 10% and TDN 60%), A1B2 (CP 10% and TDN 65%), A2B1 (CP 12% and TDN 60%), and A2B2 (CP 12% and TDN 65%).
Statistical analysis
The research data were analyzed using analysis of variance using Stastix 8 software. If significant differences in the results were obtained, the Duncan test, as outlined in Steel and Torrie (2002), was conducted.
RESULTS AND DISCUSSION
Feed consumption
The treatments showed a significant (P<0.05) interaction with dry matter (DM) consumption (Table 3). The A1B2 treatment exhibited the highest DM consumption (Figure 1), whereas the A1B1 treatment had the lowest. The difference in DM consumption was caused by differences in the protein and energy ratio between treatments. These two main nutrients play a crucial role in supporting the growth and activity of rumen microbes in degrading feed. Treatment A1B2 has a higher energy content compared to treatments A1B1 and A2B1 (Table 2). Energy is needed by rumen microbes to increase their population so it has an impact on the amount of enzyme secretion produced in the feed degradation process, which will have a direct impact on
Table 2: Nutritional composition of treatment ration.
|
Nutritional composition (%) |
Treatments |
|||
|
A1B1 |
A1B2 |
A2B1 |
A2B2 |
|
|
Dry matter |
89.59 |
90.99 |
91.71 |
90.87 |
|
Organic matter |
88.32 |
84.33 |
91.48 |
91.46 |
|
Ash |
11.67 |
15.67 |
9.51 |
8.53 |
|
Crude protein |
10.32 |
10.03 |
12.26 |
12.38 |
|
Crude fiber |
34.86 |
22.51 |
36.99 |
30.64 |
|
Crude fat |
2.17 |
2.29 |
3.30 |
3.16 |
|
Nitrogen free extract |
40.95 |
49.48 |
37.92 |
45.27 |
|
Total digestible nutrient |
59.76 |
64.73 |
59.87 |
65.03 |
|
Neutral detergent fiber |
69.76 |
57.52 |
60.86 |
65.71 |
|
Acid detergent fiber |
51.39 |
38.81 |
43.61 |
51.94 |
|
Cellulose |
31.01 |
34.99 |
28.69 |
16.72 |
|
Hemicellulose |
18.36 |
18.70 |
17.25 |
13.77 |
A1B1 (CP 10% and TDN 60%), A1B2 (CP 10% and TDN 65%), A2B1 (CP 12% and TDN 60%), and A2B2 (CP 12% and TDN 65%).
Table 3: In vivo daily consumption of treatment ration.
|
In vivo consumption (Kg/day) |
Factor A (Crude protein) |
Factor B (TDN) |
Average |
SEM |
|
|
B1 (60%) |
B2 (65%) |
||||
|
Dry matter (kg/day) |
A1 (10%) |
2.71aA±0.14 |
2.80aB±0.08 |
2.76 |
0.04 |
|
A2 (12%) |
2.69bA±0.08 |
2.60bB±0.06 |
2.65 |
||
|
Average |
2.70 |
2.71 |
|||
|
Organic matter (kg/day) |
A1 (10%) |
2.39aA±0.13 |
2.37aB±0.06 |
2.38 |
0.04 |
|
A2 (12%) |
2.43bA±0.07 |
2.38bB±0.05 |
2.41 |
||
|
Average |
2.41 |
2.37 |
|||
|
Crude protein (kg/day) |
A1 (10%) |
0.27aA±0.01 |
0.28aA±0.01 |
0.28a |
0.01 |
|
A2 (12%) |
0.33bA±0.01 |
0.32bA±0.01 |
0.33b |
||
|
Average |
0.30 |
0.30 |
|||
Different lowercase letters in the row and different uppercase letters in the same column have a significantly different effect (P<0.05).
feed digestibility. Another factor that determines the high consumption of DM is the palatability and composition of the feed ingredients that make up the ration (Arief et al., 2023a, b; Jamarun et al., 2023; Pazla et al., 2023b). The A1B2 ration is more palatable because the main energy source comes from corn, whereas in the A1B1 and A2B1 treatments, the main energy source comes from rice bran. The A2B2 treatment is lower than A1B2 because the percentage of corn used is lower than A1B1.
The A1B2 treatment contained a lower ration CP compared to A2B1 and A2B2. Diet protein is needed to provide a nitrogen source for microbes (Pazla et al., 2021b; Zain et al., 2023). However, there needs to be a balance with energy availability because balanced protein and energy will increase the rate of microbial growth. Nitrogen from protein will be used as a nitrogen source for the formation of microbial protein, whereas apart from nitrogen, the process of forming microbial protein also requires ATP, which comes from available energy. This condition will increase the rate of feed leaving the rumen (rate of passage) so that the rumen empties more quickly and stimulates livestock to consume more ration. DM consumption in the A1B2 treatment was higher than in Usman et al. (2013), namely 2.72 kg with a balance of CP 13.67% and TDN 60%.
Organic material is the largest component of dry matter rations, consisting of crude protein, crude fat, crude fiber, and extracts without nitrogen. Consumption of organic material is an indicator of energy adequacy for livestock. The treatments showed a significant (P<0.05) interaction with organic material consumption (Table 3). Treatment A1B1 showed higher OM consumption than A1B2 and A2B2, but the highest consumption was found in A2B1. The high OM consumption in A2B1 was due to the high organic material content of the ration in A2B1. The organic material content of the ration plays a very important role in determining the amount of organic material consumed (Febrina et al., 2017).
The treatments showed a significant (P<0.05) interaction with crude protein consumption (Table 3). The highest crude protein consumption was in treatment A2B1, which was not significantly different from A2B2. The high consumption of CP in this treatment was due to the high protein content of the diet. CP consumption is highly dependent on the protein content of the diet consumed by livestock (Arief and Pazla, 2023).
Digestibility of dry matter, organic matter, and crude protein
The impact of varying the ratio of CP and TDN in the treated feed had a significant (P<0.05) interaction on the OMD, DM, and CP in Pesisir cattle, as shown in Table 4. Numerically, the highest levels of DM and OM digestibility were achieved in treatment A1B2, followed by A1B1, A2B1, and A2B2. The high digestibility of DM and OM in treatment A2B1 can be attributed to the balanced values of CP and TDN in the ration. Rumen microbes require a balance of protein and energy for their activity and growth. The greater the rumen microbial population, the more food substances that can be digested (Jamarun et al., 2017a, b). Apart from that, the CP and TDN ratios of A1B2 were lower than the others. Several studies show that reducing the CP and TDN ratio results in better DM and OM digestibility. The current CP and TDN ratio of 0.25 in the feed led to decreased digestibility in male Pesisir cattle, in contrast to the ratio of 0.23 as reported by Pazla et al. (2021a). These findings align with Purbowati et al. (2008) and Nugroho et al. (2013), both of which indicated that an increase in the CP: TDN ratio harmed feed digestibility values.
The pattern of OMD is linear with DMD because it constitutes the largest component of DM. This linear pattern has been previously observed by Antonius et al. (2023); Elihasridas et al. (2023a). The organic material components in the A1B2 treatment are more easily degraded by rumen microbes, resulting in higher OMD than other treatments. Montcho et al. (2017) stated that easily soluble and degraded organic matter helps increase rumen microbial activity, enabling effective feed degradation.
Providing feed containing a balanced ration of protein and energy activates rumen microbes, which can increase the population of proteolytic and deamination bacteria, thereby enhancing feed digestibility. Efficiency in the synthesis of microbial protein is dependent on the sequential availability of ammonia, followed by the availability of energy and carbon skeletons (Febrina et al., 2016; Pazla et al., 2018a; Elihasridas et al., 2023b). In cases where ammonia becomes available more rapidly than carbohydrate fermentation, the utilization of ammonia for microbial protein formation can become inefficient. Optimal conditions for microbial protein synthesis are achieved when the availability of fermented carbohydrates aligns with the availability of the protein source (Christiyanto et al., 2005). Table 4 shows that the A1B2 showed better CP digestibility values compared to other treatments.
CP digestibility is the percentage of crude protein consumed in the diet that does not appear in feces. This digestibility is affected by the crude protein content in the feed. An increase in the crude protein content in the diet leads to higher microbial reproduction rates and a larger rumen population, which, in turn, enhances digestibility (Haryanto, 2014; Tedeschi et al., 2015). Nevertheless, an increase in protein levels in rations, when not adequately balanced with sufficient non-protein nitrogen, fails to
Table 4: In vivo digestibility of treatment ration.
|
In vivo digestibility (%) |
Factor A (Crude protein) |
Factor B (TDN) |
Average |
SEM |
|
|
B1 (60%) |
B2 (65%) |
||||
|
Dry matter |
A1 (10%) |
64.92bA±3.70 |
66.99bA±1.96 |
65.95b |
1.12 |
|
A2 (12%) |
61.34aA±2.10 |
56.15aA±1.18 |
58.75a |
||
|
Average |
63.13A |
61.57A |
|||
|
Organic matter |
A1 (10%) |
66.44bA±3.81 |
68.60bA±1.77 |
67.52b |
1.27 |
|
A2 (12%) |
62.92aA±3.46 |
58.35aA±1.09 |
60.64a |
||
|
Average |
64.68A |
63.47A |
|||
|
Crude protein |
A1 (10%) |
80.08bB±3.02 |
80.83bA±1.56 |
80.46b |
1.04 |
|
A2 (12%) |
79.45aB±2.50 |
72.18aA±1.21 |
75.82a |
||
|
Average |
79.76B |
76.51A |
|||
|
Crude Fat |
A1 (10%) |
65.88bA±2.80 |
69.72bA±1.68 |
67.80b |
1.15 |
|
A2 (12%) |
58.45aA±2.32 |
51.33aA±2.38 |
54.89a |
||
|
Average |
62.16A |
60.52A |
|||
|
Crude Fiber |
A1 (10%) |
58.92bA±3.12 |
59.83bA±3.22 |
59.87b |
1.34 |
|
A2 (12%) |
53.28aA±2.88 |
49.33aA±1.51 |
51.30a |
||
|
Average |
56.10A |
55.08A |
|||
|
Neutral detergent fiber |
A1 (10%) |
57.09bA±3.99 |
61.02bA±1.56 |
59.05b |
1.32 |
|
A2 (12%) |
51.26aA±3.69 |
45.26aA±1.33 |
48.26a |
||
|
AVERAGE |
54.18A |
53.14A |
|||
|
Acid detergent fiber |
A1 (10%) |
55.43bA±4.30 |
57.98bA±3.05 |
56.70b |
1.58 |
|
A2 (12%) |
49.43aA±4.55 |
43.97aA±0.75 |
46.70a |
||
|
Average |
52.43A |
50.97A |
|||
|
Cellulose |
A1 (10%) |
58.99bA±4.60 |
63.52bA±2.54 |
61.25b |
1.52 |
|
A2 (12%) |
52.78aA±4.09 |
47.23aA±0.96 |
50.01a |
||
|
Average |
55.88A |
55.38A |
|||
|
Hemicellulose |
A1 (10%) |
60.55bA±4.34 |
63.85bA±0.50 |
62.20b |
1.12 |
|
A2 (12%) |
54.90aA±2.99 |
50.13aA±1.11 |
52.52a |
||
|
Average |
57.73A |
56.99A |
|||
|
Extract material without nitrogen |
A1 (10%) |
68.16bA±3.38 |
71.45bA±2.19 |
69.81b |
1.03 |
|
A2 (12%) |
63.99aA±1.72 |
57.72aA±0.94 |
60.86a |
||
|
Average |
66.08A |
64.59A |
|||
|
Total digestible nutrient |
A1 (10%) |
65.44bA±1.05 |
67.39bA±1.69 |
66.42b |
0.74 |
|
A2 (12%) |
61.64aA±2.45 |
56.07aA±0.76 |
58.85a |
||
|
Average |
63.54A |
61.73A |
|||
Different lowercase letters in the row and different uppercase letters in the same column have a significantly different effect (P<0.05)
promote microbial growth in the rumen. Similarly, Teti et al. (2018) noted that achieving a balance between protein and total digestible nutrients (TDN) is crucial for optimal fermentation efficiency. In line with this, Rosmalia et al. (2022a) emphasized that the addition of protein sources alone may not stimulate rumen microbial growth without the supplementation of soluble carbohydrates. Rosmalia et al. (2022b) pointed out that rations with higher TDN content typically undergo more extensive fermentation compared to those with lower TDN content. Moradi and Zadeh (2013) indicated that achieving synchronization or balance between the availability of energy and protein in the rumen can boost microbial activity and microbial protein synthesis in the rumen. In the current study, the A1B2 treatment produced better digestibility compared to the A1B1, A2B1, and A2B2 treatments.
Extract ether digestibility (EED)
The treatments revealed a significant (P<0.05) interaction regarding crude fat digestibility, with the highest value found in treatment A1B2 and the lowest in A2B2. In Table 4, it is evident that fat digestibility tends to decrease with increasing protein with the same TDN content. However, fat digestibility tends to increase with raising TDN content with the same protein level. A high crude fat concentration in the A2B2 treatment diet led to decreased fat digestibility, whereas a low crude fat concentration in the A1B1 treatment resulted in increased crude fat digestibility. The crude fat content in the treatment influences the level of crude fat digestibility. As shown in Table 1, treatment A2B2 has the highest fat content among the other treatments, resulting in the lowest fat digestibility. The substantial use of palm kernel cake in A2B1 and A2B2 impacts the level of crude fat digestibility. This aligns with Polii et al. (2020) and Makmur et al. (2019) who stated that high crude fat content in rations reduces nutritional digestibility. Alhadas et al. (2023) also reported that an increase in protein levels and a decrease in NFC (Non-fiber Carbohydrate) levels in low-fat DDGS-based rations and high-protein concentrates influenced a decrease in crude fat digestibility due to the higher crude fat composition in the treatment rations.
The reduction in fat digestibility was correlated with a decrease in the digestibility of DM and OM, as demonstrated in Table 4. There was a decrease in DDM and DOM in A2B2. The decrease in fat digestibility is caused by an imbalance in the concentration of protein and energy in the ration, resulting in microbes experiencing deficiencies in their body’s protein synthesis, which impairs feed digestion activities. The recent study explained that a ration with a 10% CP and a 60-65% TDN demonstrates an optimal balance of protein and energy for increasing fat digestibility. In line with Han et al. (2022), who reported that the balance of protein and energy impacts crude fat digestibility. A balance of protein and energy capable of optimizing the protein formation process in rumen microbial bodies will enhance the productivity of rumen microbes in digesting feed (Putri et al., 2019, 2021; Sari et al., 2022; Zain et al., 2020).
There was a significant (P<0.05)interaction between protein and energy levels on extract ether digestibility (EED). The highest EED was observed in the A1B2 treatment, while the lowest was in the A2B2 treatment. As shown in Table 4, an increase in protein levels at the same TDN level tended to decrease EED. Conversely, an increase in TDN levels at the same protein level tended to increase EED. It can be explained that a high concentration of extract ether in A2B2 depressed its digestibility, while the lowest concentration of extract ether in A1B1 improved its digestibility. The composition of extract ether in the experimental ration affected EED. Table 2 showed that A2B2 consisted of 3.16% extract ether, which is higher than in other treatments, resulting in the lowest EED value. The higher composition of palm kernel cake in the ration, especially in A2B1 and A2B2 was associated with a decrease in EED. This aligns with Polii et al. (2020), who noted that an elevated content of ether extract in the diet contributes to a reduction in nutrient digestibility. Alhadas et al. (2023) also reported that an increase in protein levels and a decrease in NFC (Non-fiber Carbohydrate) levels in low-fat dried distillers grains without soluble (DDGS) and high-concentrate rations affected the decrease in extract ether digestibility due to an increase in ether extract composition in the ration.
In addition to being influenced by extract ether composition, EED is associated with a decrease in DMD and OMD. As seen in Table 4, the decrease in DMD and OMD corresponds with the decrease in EED. This correlation can be explained by the fact that dry matter consists of all nutrients, while OM consists of all nutrients except ash. Furthermore, the decrease in EED was attributed to the lack of synchronization between protein and energy in the rumen, leading to a deficiency in microbe protein synthesis. This, in turn, results in a reduction in digestibility activities in the rumen. The recent study explains that a ration consisting of 10% protein and 60-65% TDN demonstrated the optimal protein and energy ratio that can increase EED. This is in line with Han et al. (2022), who reported that the balance of protein and energy influences the increase in ether extract digestibility. The protein and energy ratio can optimize microbe protein synthesis. The efficiency of nutrient digestibility is influenced by a high population of rumen microbes, as indicated by Putri et al. (2019, 2021), Sari et al. (2022), and Zain et al. (2020).
Table 5: Body weight gain of treatment ration.
|
Parameter |
Factor A (Crude protein) |
Factor B (TDN) |
Average |
SEM |
|
|
B1 (60%) |
B2 (65%) |
||||
|
Body weight gain (Kg/day) |
A1 (10%) |
0.49±0.14 |
0.52±0.16 |
0.50 |
00.06 |
|
A2 (12%) |
0.47±0.09 |
0.44±0.06 |
0.46 |
||
|
Average |
0.48 |
0.48 |
|||
Nitrogen free extract digestibility (NFED)
The levels of protein and energy in the diet significantly (P<0.05) influenced the digestibility of NFE, with the highest digestibility value found in the A1B2 treatment and the lowest in the A2B2 treatment. It showed that the A1B2 is more synchronized than the A2B2. In Table 6, it can be seen that increasing the TDN level at the same protein level tends to increase nitrogen-free extract (NFE) digestibility, although not significantly. Meanwhile, increasing the protein level at the same TDN level results in the NFE digestibility value tending to decrease significantly (P<0.05). It indicates that an increase in protein levels without a TDN balance results in protein and energy asymmetry in the protein formation process of the microbial body, reducing the ability of the microbes to digest food. Increasing protein levels will not stimulate microbial growth without a balanced energy source (Sahroni et al., 2021). However, adding the TDN level to a protein level of 10% showed the highest NFE digestibility (10% and TDN of 60-65%). This showed that 65% of TDN has met the energy needs of livestock and rumen microbes. The most dominant components of NFE are non-structural carbohydrates, such as starch and monosaccharides. This component is found in many cereal plants, including corn. The corn composition of 25% in A1B2 contributed to the high digestibility of NFE compared to other treatments. In line with Wiryawan et al. (2017), which states that NFE with high starch content will contribute to a high energy source. This energy will be used by rumen microbes in the body’s protein synthesis process. Increasing the efficiency of microbial protein formation will increase nutrient digestibility (Pazla et al., 2018a; Putri et al., 2021).
Table 6: Ration efficiency of treatment ration.
|
Parameter |
Factor A (Crude protein) |
Factor B (TDN) |
Average |
SEM |
|
|
B1 (60%) |
B2 (65%) |
||||
|
Ration efficiency (%) |
A1 (10%) |
18.33±0.73 |
18.59±2.10 |
18.46 |
0.98 |
|
A2 (12%) |
17.61±3.05 |
17.02±2.00 |
17.32 |
||
|
Average |
17.96 |
17.81 |
|||
The levels of protein and energy in the experimental ration significantly (P<0.05) influenced the digestibility of NFED, with the highest value found in the A1B2 treatment and the lowest in the A2B2 treatment. This demonstrates that a ration consisting of 10% protein and 65% TDN is more synchronous than a ration with 12% protein and 65% TDN. In Table 6, it can be observed that increasing TDN levels at the same protein level tends to increase NFED, although not significantly. On the other hand, increasing protein levels at the same TDN level results in a significant (P<0.05) decrease in NFED. This indicates that an increase in protein levels without a balanced TDN source results in unsynchronized protein and energy levels. This imbalance interferes with microbial protein synthesis, reducing the ability of microbes to digest feed. An increase in protein levels alone will not be sufficient to stimulate microbial growth without a balanced energy source (Sahroni et al., 2021). However, increasing TDN levels to 60-65% at a protein level of 10% showed the highest NFED. This suggested that a TDN level of 65% meets the needs of cattle and rumen microbes. The most dominant component of NFE is non-structural carbohydrates, such as starch and monosaccharides, which are found in many cereal plants, including corn. The corn composition in A1B2 contributed to the high NFED compared to other treatments. This aligns with Wiryawan et al. (2017) who stated that NFE with a high starch content provides a significant energy source. This energy is utilized by rumen microbes in the process of protein synthesis, ultimately enhancing nutrient digestibility (Pazla et al., 2018a; Putri et al., 2021).
Total digestible nutrient (TDN)
Total digestible nutrient is the energy that can be digested and utilized by livestock, obtained from the sum of the digestibility of protein, crude fiber, NFE, and crude fat. The research results showed that the levels of protein and TDN in the ration influence the amount of TDN that can be utilized by livestock. We demonstrated that increasing protein and TDN levels significantly (P<0.05) affected TDN digestibility. Increasing TDN levels at the same protein level tended to increase TDN digestibility, although not significantly. The increase in digestibility in A1B2 was due to the higher TDN content. In line with Teti et al. (2018), who stated that a high TDN content in the ration is more easily degraded by rumen microbes. In contrast, increasing the protein level at the same TDN level resulted in a significant (P<0.05) decrease in TDN digestibility. This explains that the increase in protein levels is not balanced with the TDN content, resulting in suboptimal nutrient absorption. Consistency in the synchronization between protein and energy levels will increase microbial protein synthesis. Energy that degrades easily must be balanced with protein that degrades easily, ensuring that no nutrients go to waste (Syamsi et al., 2022).
Total digestible nutrient represents the energy that livestock can digest and utilize, derived from the summation of protein, crude fiber, NFE (Nitrogen-Free Extract), and crude fat digestibility. The current research has demonstrated a significant (P<0.05) influence of protein and TDN levels on TDN utilization in livestock. An increase in TDN levels at the same protein level tended to enhance TDN digestibility, although not statistically significant. The observed improvement in digestibility in A1B2 can be attributed to the increased TDN content. This finding aligns with the study conducted by Teti et al. (2018), which emphasized that rations with higher TDN content are more readily degraded by rumen microbes. Conversely, raising the protein level at a consistent TDN level resulted in a significant (P<0.05) decrease in TDN digestibility. This discrepancy suggests that the elevated protein levels lack balance with the TDN content, leading to suboptimal nutrient absorption. Achieving a harmonious synchronization between protein and energy levels is crucial for enhancing microbial protein synthesis. This ensures that easily degradable energy complements readily degradable protein, minimizing nutrient wastage (Syamsi et al., 2022).
Crude fiber digestibility (CFD)
Crude fiber plays a pivotal role as an energy source for rumen microbes and serves as a rumen filler for ruminants. We demonstrated that variations in the combination of protein and energy levels in the ration lead to significant (P<0.05)differences in crude fiber digestibility levels. As depicted in Table 4, there is a trend towards increased crude fiber digestibility with higher TDN levels at consistent protein levels, even though this difference is not statistically significant. Conversely, when keeping the protein level at 12%, there is a tendency toward decreased crude fiber digestibility, although not statistically significant. However, significantly (P<0.05), varying the protein level in diets with the same TDN level results in a notable decrease in crude fiber digestibility.
This observation aligns with previous studies that report increased TDN levels lead to enhanced crude fiber digestibility, while higher protein levels tend to decrease it (Teti et al., 2018). Zhou et al. (2015) reported that elevating energy levels at consistent protein levels positively impacts crude fiber digestibility. This phenomenon can be explained by the fact that 65% of TDN meets the energy requirements of cattle, with further enhancement achievable through balanced protein levels. The level of feed digestibility, including crude fiber, is significantly influenced by the composition of the feed ration. As the crude fiber content in the ration increases, digestibility tends to decrease. The observed reduction in crude fiber digestibility in A2B1 and A2B2 can be attributed to the high crude fiber content in these rations.
Crude fiber fraction digestibility
Fiber is found in many cell walls and comprises the main components of hemicellulose, cellulose, and lignin. Cell walls that are insoluble in neutral detergents are referred to as NDF, consisting of lignin, cellulose, hemicellulose, and small amounts of protein, nitrogen bonds, minerals, and cuticle. NDF plays a crucial role for ruminants as it is closely linked to dry matter intake. This study reports a significant (P<0.05) influence of protein and energy balance on NDF digestibility. The highest NDF value was observed in treatment A1B2, while the lowest was in A2B2. Furthermore, the highest ADF value was found in A1B2, and the lowest was in A2B2. In line with these findings, the highest digestibility values for cellulose and hemicellulose were recorded in A1B2, and the lowest values were in A2B2 (Table 4). These results indicated that A1B2 offers the most optimal protein and energy ratio for enhancing digestibility. Increasing the TDN level at the same protein level tended to enhance the digestibility of the fiber fraction, although this difference was not statistically significant. Conversely, increasing the protein level at the same TDN level resulted in a significant (P<0.05) decrease in the digestibility of the fiber fraction. This suggests that TDN levels of up to 65% at 10% protein levels provide an optimal balance of protein and energy. The availability of nitrogen and protein degradation, in conjunction with VFA production from energy degradation, fosters favorable conditions for microbial protein synthesis. Microbes become more active in degrading the fiber fraction, in agreement with Emmanuel et al. (2015), who emphasized that a balanced availability of nitrogen and soluble energy sources enhances feed fermentation in the rumen.
Body weight gain, efficiency, and ration conversion
The findings from the study regarding the average daily body weight gain, feed efficiency, and feed conversion ratio of the experimental diets given to Pesisir heifers are displayed in Tables 5, 6, and 7. It is evident from Table 5 and Figure 2 that the A1B2 treatment achieved the highest average daily body weight gain, although no significant interaction was observed compared to the other treatments. This weight gain is accompanied by high DM consumption, also observed in A1B2. The lowest body weight gain is found in treatment A2B2. Animal body weight is consistently directly proportional to the level of feed consumption. Increased feed consumption leads to greater body weight, and this also affects population growth, as indicated by Ningrat et al. (2020) and Malindo et al. (2023). Weight gain occurs when the quantity of food consumed exceeds the animal’s requirements, and the surplus nutrients are converted into meat and fat tissue increasing body weight. Ningrat et al. (2018) demonstrated that when the amount of feed consumed is insufficient to meet the livestock’s needs, it results in a decrease in body weight.
Table 7: Feed conversion ratio of treatment ration.
|
Parameter |
Factor A (Crude protein) |
Factor B (TDN) |
Average |
SEM |
|
|
B1 (60%) |
B2 (65%) |
||||
|
Feed conversion ratio |
A1 (10%) |
5.53±0.73 |
5.38±2.10 |
5.46 |
0.98 |
|
A2 (12%) |
5.72±3.05 |
5.91±2.00 |
5.32 |
||
|
Average |
5.96 |
5.81 |
|||
The body weight gain was higher than the body weight gain of male Pesisir cattle weaned until the age of 2 and 3-4 years. The body weight gain was less than what was reported by Afdal and Khasrad (2006), who recorded a PBB range of 0.58-0.80 kg/head/day for Pesisir cattle. The higher PBB value in their study was attributed to the ration provided, which had a TDN value of 68%, and the experimental livestock experienced a compensatory growth effect.
In A2B2, the weight gain was at its minimum. The conversion rate for the A2B2 diet indicated that to achieve an increase of 1 kg in body weight per day, 5.91 kg of dry ration material is necessary (Table 7). It can be concluded that providing the A2B2 treatment does not result in efficient feed conversion. Therefore, the feed conversion in the A2B2 treatment is suboptimal due to a significant amount of feed being consumed with less efficient meat production. However, this combination did not have a significant impact on rumen microbes. Rumen microbes require balanced CP and TDN values for their growth. In alignment with this, Pazla et al. (2018b) have also illustrated that the quality of the feed, particularly its protein, energy, and crude fiber content, significantly influences livestock’s capacity to convert feed into meat.
The treatment with the lowest feed conversion rate was A1B2. An effective feed conversion rate is achieved when modest feed consumption leads to a substantial increase in body weight. This is likely because the feed consumption in treatment A1B2 meets the physiological requirements of the livestock with an optimal balance of protein and energy content. It has been noted by Pazla et al. (2021a) that a more favorable conversion ratio is achieved when there is a higher nutritional value in the ration.
The feed conversion values in treatments A1B1 and A2B1 were suboptimal. The results indicated that ration conversion in treatment A1B2 was lower than in the other treatments. This implies that the presence of energy sources in treatment A1B2 is more efficient, enabling rumen microorganisms to operate with greater efficiency.
The best feed conversion was achieved in A1B2. This situation demonstrates that the A1B2 ration is of excellent quality for livestock, as it exhibits high palatability. As noted by Suyitman et al. (2020), feed quality plays a significant role in determining feed conversion. Additionally, high-quality feed leads to higher Average Daily Gain (ADG). Feed conversion is influenced by factors such as the type of animal, genetics, the cow’s condition, feed palatability, seasonal conditions, age, management, the animal’s ability to digest feed, types of feed ingredients, nutrient availability in the ration, and ADG (Pazla et al., 2018b; Suyitman et al., 2020; Jusman et al., 2020). According to Sugiharto et al. (2004), cattle fed with local feed, following the practices of dryland breeders, attain a feed conversion ratio of 13.6. This is in contrast to the ideal feed conversion for beef cattle, which is 9, as suggested by Tillman et al. (2005).
Treatment A1B2 demonstrated the best feed efficiency, resulting in higher body weight gain in this treatment compared to treatments A1B1, A2B1, and A2B2 (Table 6). The optimal balance of protein and energy in treatment A1B2 had a positive influence on the performance of rumen microbes in breaking down feed, ultimately leading to increased feed efficiency, as reported by Pazla et al. (2021a). The lower feed efficiency in treatments A1B1, A2B1, and A2B2 was attributed to an imbalance in crude protein and TDN content in the ration. Following Ariesafera (2019), who emphasized the close relationship between the CP and TDN ratio in rations and feeding efficiency. Efficient feed consumption in Pesisir cattle is achieved when there is a well-balanced ratio of CP and TDN in their diet. The feed efficiency observed exceeded that reported by Jusman et al. (2020), which was 7.78%, and that of Suyitman et al. (2020), which was 12.22%. This variation can be attributed to factors such as the cattle type, the composition of feed ingredients in the diet, and the quality of the feed used.
Estrous percentage
Figure 3 shows that treatment A1B2 achieved the highest estrous percentage. The speed of estrous in cows is greatly influenced by both the quantity and the nutritional quality of the ration provided. Treatment A2B1 had the highest dry matter consumption (Table 3), along with the highest level of digestibility (Table 4). Optimal nutrients consumed and utilized by the livestock’s body stimulate the production of reproductive hormones, leading to quicker estrous. Conversely, when ration consumption and nutrient digestibility are low, it slows down the process of entering estrous due to insufficient nutrients to stimulate the formation of reproductive hormones. Inadequate feed consumption directly affects protein intake, which plays a vital role in hormone production, including reproductive hormones. Insufficient protein leads to a shortage of reproductive hormones, resulting in reproductive disorders such as silent heat and ovarian issues. Yendraliza (2013) noted that reduced food levels generally delay the onset of puberty, while high food levels can accelerate it and increase body weight. Wiltbank et al. (2006) added that rapid weight gain between birth and weaning, as well as between weaning and 396 days of age, speeds up the onset of puberty in heifers. Delayed puberty due to insufficient food may result from low levels of gonadotropins produced by the adenohypophyseal gland, a lack of ovarian response, or possibly the ovaries’ failure to produce sufficient amounts of estrogen (Yendraliza, 2013). Dairy cows demonstrate that this combination can delay sexual maturity and suppress the signs of heat (Salisbury and Vandemark, 1985). In the ovaries, low feed intake delays puberty, accompanied by a decrease in the development of ovarian follicles, so low feed intake in female cows can cause the dominant follicle to remain smaller (Bergfeld et al., 1994).
CONCLUSION
Providing a ration containing 10% CP and 65% TDN increased consumption, feed digestibility, production performance, and the estrous percentage in Pesisir heifers.
ACKNOWLEDGMENTS
This research was funded by the Indonesian National Research Agency under contract numbers 88/IV/ KS/11/2022 and T/283/UN16.17/PT.01.03/KS/2022. We would like to express our gratitude to the Center for Breeding Superior Livestock and Forage for Animal Feed Padang Mengatasand Tuah Sakato Artificial Insemination Center in Payakumbuh. Additionally, we extend our appreciation to the laboratory assistants in ruminant nutrition at the Faculty of Animal Husbandry, Andalas University, for their invaluable support in conducting laboratory analyses.
NOVELTY STATEMENT
We identified the ideal protein to energy ratio for enhancing nutrient intake, production performance, nutrient digestibility, and the percentage of first estrous in Pesisir cattle. This research represents a novel contribution, as no previous studies have investigated this aspect.
AUTHOR’S CONTRIBUTION
Conceptualization: MZ, ZU, TT, WN, J and FA. Data curation: RP, MZ, FA, and EMP. Formal analysis: FA, RP, and EMP. Funding acquisition: MZ and FA. Methodology: FA, YM, H, and M. Project administration: MM and EMP. Supervision: MZ and ZU. Validation: MZ, FA, and RP. Writing-original draft: RP, FA, and EMP. Writing-review and editing: RP.
Ethical approval
All research procedures carried out have been approved and monitored by the Research Ethics Committee of the Faculty of Medicine, Andalas University, Padang, West Sumatra with the following numbers 297/UN.16.2/KEP-FK/2023. The conducted in-vivo trial also followed research ethics related to livestock as outlined in Section 66 of the Republic of Indonesia Government Law Number 18 of 2009 concerning the management, breeding, euthanasia, and appropriate treatment and welfare of animals. We followed the guidelines provided in the ‘Guide for the Care and Use of Agricultural Animals in Research and Teaching’ outlined by the Federation of Animal Science Societies (American Dairy Science Association, 2020).
Conflict of interest
The authors have declared no conflict of interest.
References
Afdal M, Khasrad (2006). Effect of concentrate level and the length of fattening on the growth, feed intake and feed conversion efficiency of Pesisir cows. The 4th ISTAP. pp. 335-338.
Alhadas HM, Valadares Filho SC, Tedeschi LO, Vilela RSR, Souza GAP, Lage SM, Oliveira KR, Pacheco MVC, Pucetti P, Silva JT, Godoi LA, Silva FAS, Rennó LN, Paulino MF (2023). Effects of low-fat dried distillers grains on nutrient intake and digestibility in high-concentrate diets. Livest. Sci., 269: 1–8. https://doi.org/10.1016/j.livsci.2023.105167
American Dairy Science Association, American Society of Animal Science and Poultry Science Association (2020). Guide for the Care and Use of Agricultural Animals in Research and Teaching Fourth edition. www.adsa.org
Antonius, Pazla R, Putri EM, Negara W, Laia N, Ridla M, Suharti S, Jayanegara A, Asmairicen S, Marlina L, Marta Y (2023). Effectiveness of herbal plants on rumen fermentation, methane gas emissions, in vitro nutrient digestibility, and population of protozoa. Vet. World, 16(7): 1477-1488. https://doi.org/10.14202/vetworld.2023.1477-1488
Arief, Pazla R (2023). Milk production and quality of etawa crossbred goats with non-conventional forages and palm concentrates. Am. J. Anim. Vet. Sci., 18(1): 9-18. https://doi.org/10.3844/ajavsp.2023.9.18
Arief, Pazla R, Jamarun N, Rizqan (2023a). Production performance, feed intake and nutrient digestibility of Etawa crossbreed dairy goats fed tithonia (Tithonia diversifolia), cassava leaves and palm kernel cake concentrate. Int. J. Vet. Sci., 12(3): 428-435. https://doi.org/10.47278/journal.ijvs/2022.211
Arief, Pazla R, Rizqan, Jamarun N (2023b). Influence of Tithonia diversifolia, cassava and palm concentrate combinations on milk production and traits in etawa crossbred. Adv. Anim. Vet. Sci., 11(4): 568-577. https://doi.org/10.17582/journal.aavs/2023/11.4.568.577
Ariesafera A (2019). Pengaruh pemberian level TDN (Energi) dan protein kasar silase ransum komplit berbasis limbah tebu dan limbah kubis terhadap keuntungan usaha penggemukan Kerbau. Thesis, Andalas University, Padang, Indonesia.
Association of Official Agricultural Chemists (2016). Official methods of analysis of AOAC International. 20th edn, 2016, ed. Dr George W Latimer, Jr. USA.
Basyar B (2021). Beef cattle farm development policies to overcome beef distribution problem in Indonesia: A literature review. Am. J. Anim. Vet. Sci. 16(1): 71-76. https://doi.org/10.3844/ajavsp.2021.71.76
Bergfeld EG, Kojima FN, Cupp AS, Wehrman ME, Peters KE, Garcia-Winder M, Kinder JE (1994). Ovarian follicular by level of dietary energy intake. Biol. Reprod., 51(5): 1051-1057. https://doi.org/10.1095/biolreprod51.5.1051
BPS (2020). Jumlah sapi pesisir. Badan Pusat Statistik Indonesia.
Christiyanto M, Soejono M, Utomo R, Hartadi H, Widyobroto BP (2005). The nutrient digestibility of different proteinenergy precursor rations in dairy cattle fed on a basal diet of king grass. J. Trop. Anim. Agric., 30(4): 242-247.
Elihasridas, Zain M, Ningrat RWS, Erpomen, Putri EM, Makmur M (2023a). In vitro nutrient digestibility and ruminal fermentation characteristics of ammoniated and fermented treatment of soaked and unsoaked Cymbopogon nardus waste. Int. J. Vet. Sci., 12(3): 395-400. https://doi.org/10.47278/journal.ijvs/2022.204
Elihasridas, Zain M, Pazla R, Sowmen S, Aini Q (2023b). In-vitro digestibility of ammoniated aromatic supplemented lemongrass waste. Adv. Anim. Vet. Sci., 11(8): 1368-1376. https://doi.org/10.17582/journal.aavs/2023/11.8.1368.1376
Emmanuel N, Patil NV, Bhagwat SR, Lateef A, Xu K, Liu H (2015). Effects of different levels of urea supplementation on nutrient intake and growth performance in growing camels fed roughage based complete pellet diets. Anim. Nutr., 1(4): 356–361. https://doi.org/10.1016/j.aninu.2015.12.004
Febrina D, Jamarun N, Zain M, Khasrad (2016). The effects of P, S and Mg supplementation oil palm fronds fermented by Phanerochayte crysosporium on rumen fluid characteristics and microbial protein synthesis. Pak. J. Nutr., 15(3): 299-304. https://doi.org/10.3923/pjn.2016.299.304
Febrina D, Jamarun N, Zain M, Khasrad (2017). Effects of using different levels of oil Palm Fronds (FOPFS) fermented with Phanerochaete chrysosporium plus minerals (P, S and Mg) instead of Napier grass on nutrient intake and the growth performance of goats. Pak. J. Nutr., 16(8): 612-617. https://doi.org/10.3923/pjn.2017.612.617
Han F, Wang J, Chen L, Zhong W (2022). Effects of dietary protein and energy levels on growth performance, nutrient digestibility, and serum biochemical parameters of growing male mink (neovison vison). Front. Vet. Sci., 9: 1–9. https://doi.org/10.3389/fvets.2022.961461
Hartatik T, Putra DE, Volkandari SD, Kanazawaand T, Sumadi (2018). Genotype analysis of partial growth hormone gene (GH891│MspI) in Pesisir cattle and Simmental-Pesisir crossbred cattle. J. Ind. Trop. Anim. Agric., 43(1): 1-8. https://doi.org/10.14710/jitaa.43.1.1-8
Haryanto B (2014). Manipulating protein degradability in the rumen to support higher ruminant production. Wart, 24(3): 131-138. https://doi.org/10.14334/wartazoa.v24i3.1070
Hendri Y (2013). Development of Pesisir cattle as a local breed of West Sumatera. J. Litbang Pertanian, 32: 39-45.
Jamarun N, Pazla R, Arief, Elihasridas, Yanti G, Sari RWW, Ikhlas Z (2023). The impact of mangrove (Rhizophora apiculata) leaves hay and fermented Tithonia diversifolia on intake, nutrient digestibility and body weight gain of goat. Adv. Anim. Vet. Sci., 11(9): 1441-1450. https://doi.org/10.17582/journal.aavs/2023/11.9.1441.1450
Jamarun N, Zain M, Arief, Pazla R (2017a). Effects of calcium (Ca), phosphorus (P) and manganese (Mn) supplementation during oil palm frond fermentation by Phanerochaete chrysosporium on rumen fluid characteristics and microbial protein synthesis. Pak. J. Nutr., 16(6): 393-399. https://doi.org/10.3923/pjn.2017.393.399
Jamarun N, Zain M, Arief, Pazla R (2017b). Populations of rumen microbes and the in vitro digestibility of fermented oil palm fronds in combination with Tithonia (Tithonia diversifolia) and elephant grass (Pennisetum purpureum). Pak. J. Nutr., 17(1): 39-45. https://doi.org/10.3923/pjn.2018.39.45
Jusman V, Arnim, Marlida Y, Pazla R (2020). Production performance and Ongole crossbred cattle receiving Saccharomyces cerevisiae supplementation. J. Res. Agric. Anim. Sci., 7: 53-58.
Makmur M, Zain M, Marlida Y, Khasrad and Jayanegara A (2019). Fatty acids composition and biohydrogenation reduction agents of tropical forages. Biodiversitas, 20(7): 1917-1922. https://doi.org/10.13057/biodiv/d200718
Malindo R, Wicaksono HAD, Jayanegara A, Sjofjan O, Chuzaemi S (2023). Sustainable feed supply for bali cattle breeding center in pulukan, bali,indonesia: A system dynamics modelling. Adv. Anim. Vet. Sci., 11(7): 1209-1217. https://doi.org/10.17582/journal.aavs/2023/11.7.1209.1217
Montcho M, Babatounde S, André A, Guy M, Bougouma Y, Valerie (2017). In vitro digestibility and fermentation kinetics of agricultural and agro-industrial by-products used in ruminant feeding in Benin Republic. J. Appl. Biol. 116(1): 28-41. https://doi.org/10.4314/jab.v116i1.11
Moradi KN, Zadeh JB (2013). Synchronization of energy and protein on supply synthesis microbial protein. Int. J. Adv. Bio. Biol. Res. 1(6): 594-600. http://www.ijabbr.com/article_7778_8e1f53f4e861d2720e552cdd55d22619.pdf
Ningrat RWS, Zain M, Elihasridas, Makmur M, Putri EM, Sari YC (2020). Effect of dietary supplementation based on ammoniated palm frond with saccharomyces cerevisiae and gambier leaves waste on nutrient intake and digestibility, daily gain, and methane production of Simmental cattle. Adv. Anim. Vet. Res., 8(12): 1325-1332. https://doi.org/10.17582/journal.aavs/2020/8.12.1325.1332
Ningrat RWS, Zain M, Erpomen, Suryani H (2018). Effects of supplementation of different sources of tannins on nutrient digestibility, methane production, and daily weight gain of beef cattle fed an ammoniated oil palm frond-based diet. Int. J. Zool. Res., 14(1): 8-13. https://doi.org/10.3923/ijzr.2018.8.13
Nugroho D, Purnomoadi A, Riyanto E (2013). The effects of different ratios of dietary crude protein and total digestible nutrients on the energy utilization of native sheep. Sai. Pet., 11(2): 63-69. https://doi.org/10.20961/sainspet.11.2.63-69
Pazla R, Adrizal, Sriagtula R (2021a). Intake, nutrient digestibility and production performance of Pesisir cattle fed tithonia diversifolia and Calliandra calothyrsus-based rations with different protein and energy ratios. Adv. Anim. Vet. Res., 9(10): 1608–1615. https://doi.org/10.17582/journal.aavs/2021/9.10.1608.1615
Pazla R, Jamarun N, Elihasridas, Arief, Yanti G, Ikhlas Z (2023b). The impact of replacement of concentrates with fermented tithonia (Tithonia diversifolia) and avocado waste (Persea americana miller) in fermented sugarcane shoots (Saccharum officinarum) based rations on consumption, digestibility, and production performance of Kacang goat. Adv. Anim. Vet. Sci., 11(3): 394-403. https://doi.org/10.17582/journal.aavs/2023/11.3.394.403
Pazla R, Jamarun N, Zain M, Arief (2018a). Microbial protein synthesis and in vitro fermentability of fermented oil palm fronds by Phanerochaete chrysosporium in combination with tithonia (Tithonia diversifolia) and elephant grass (Pennisetum purpureum). Pak. J. Nutr., 17(10): 462-470. https://doi.org/10.3923/pjn.2018.462.470
Pazla R, Jamarun N, Zain M, Yanti G and Chandra RH (2021b). Quality evaluation of Tithonia (Tithonia diversifolia) with fermentation using Lactobacillus plantarum and Aspergillus ficuum at different incubation times. Biodiversitas, 22(9): 3936-3942. https://doi.org/10.13057/biodiv/d220940
Pazla R, Zain M, Despal, Tanuwiria UH, Putri EM, Makmur M, Zahera R, Sari LA, Afnan IM, Rosmalia A, Yulianti YI, Putri SD, Mushawwir A, Apriliana RA (2023a). Evaluation of rumen degradable protein values from various tropical foliages using in vitro and in situ methods. Int. J. Vet. Sci., 12(6): 860-868.
Pazla R, Zain M, Ryanto I, Dona A (2018b). Supplementation of minerals (phosphorus and sulfur) and Saccharomyces cerevisiae in a sheep diet based on a cocoa by product. Pak. J. Nutr., 17(7): 329–335. https://doi.org/10.3923/pjn.2018.329.335
Pighin D, Pazos A, Chamorro V, Paschetta F, Cunzolo S, Godoy F, Messina V, Pordomingo A, Grigioni A (2016). A contribution of beef to human health: A review of the role of the animal production systems. Sci. World J., 2016: 1-10. https://doi.org/10.1155/2016/8681491
Polii DN, Waani M, Pendong A (2020). Kecernaan protein Kasar dan Lemak Kasar pada Sapi Perah Peranakan FH (Frisian Holstein) yang diberi Pakan Lengkap Berbasis Tebon Jagung. J. Zool., 40(2): 482–492. https://doi.org/10.35792/zot.40.2.2020.28632
Purbowati E, Sutrisno CI, Baliarti E, Budhi SPS, Lestariana W (2008). Complete feed energy utilization protein-energy content is different on local fattened male sheep on a feedlot basis. J. Pen. Pet. Trop., 33(1): 59-65.
Putri EM, Zain M, Warly L, Hermon H (2019). In vitro evaluation of ruminant feed from West Sumatera based on chemical composition and content of rumen degradable and rumen undegradable proteins. Vet. World, 12(9): 1478–1483. https://doi.org/10.14202/vetworld.2019.1478-1483
Putri EM, Zain M, Warly L, Hermon H (2021). Effects of rumen-degradable-to-undegradable protein ratio in ruminant diet on in vitro digestibility, rumen fermentation, and microbial protein synthesis. Vet. World. 14(3): 640–648. https://doi.org/10.14202/vetworld.2021.640-648
Rosmalia A, Permana I, Despal (2022a). Synchronization of rumen degradable protein with non-fiber carbohydrate on microbial protein synthesis and dairy ration digestibility. Vet. World, 15(2): 252-261. https://doi.org/10.14202/vetworld.2022.252-261
Rosmalia A, Permana I, Dewi NA, Despal (2022). Reformulation of Dairy cattle concentrate based on rumen degradable protein to undegradable protein ratio at different energy levels: In vitro study. IOP Conf. Ser. Earth Environ. Sci., 1020: 012008. https://doi.org/10.1088/1755-1315/1020/1/012008
Sahroni WP, Permana IG, Despal (2021). Reformulation of dairy cow diets based on rumen degradable protein and total digestible nutrient with varying levels on in vitro fermentability and digestibility. IOP Conf. Ser.: Earth Environ. Sci., 888(1): 0–9. https://doi.org/10.1088/1755-1315/888/1/012075
Salisbury GW, dan Vandemark MIL (1985). Fisiologi dan Inseminasi Buatan Pada Sapi. (diterjemah oleh R. Djanuar). UGM Press. Yogyakarta.
Sari RM, Zain M, Jamarun N, Ningrat RWS, Elihasridas, Putri EM (2022). Improving rumen fermentation characteristics and nutrient digestibility by increasing rumen degradable protein in ruminant feed using Thitonia diversifolia and Leucaena leucocephala. Int. J. Vet. Sci., 11(3): 353–360. https://doi.org/10.47278/journal.ijvs/2021.121
Steel RGD, Torrie JH (2002). Principle, and procedures of statistics: A biometrical approach. 3rd Ed. McGraw Hill Book, New York.
Sugiharto Y, Ngadiyono N, Basuki P (2004). Produktfitas sapi peranakan ongole pada pola pemeliharaan system perkampungan ternak dan kandang individu di Kabupaten Bantul. J. Agrosains, 17(2): 191-202.
Suyitman, Warly L, Rahmat A, Pazla R (2020). Digestibility and performance of beef cattle fed ammoniated palm leaves and fronds supplemented with minerals, cassava leaf meal, and their combinations. Adv. Anim. Vet. Sci., 8(9): 91–96. https://doi.org/10.17582/journal.aavs/2020/8.9.991.996
Syamsi AN, Ifani M, Subagyo Y (2022). The protein-energy synchronization index of the tropical legumes for ruminants. J. Pet., 19(1): 29–37. https://doi.org/10.24014/jupet.v19i1.15415
Tedeschi LO, Fox DG, Fonseca MA, Cavalcanti LVL (2015). Models of protein and amino acid requirements for cattle. R. Bras. Zootec., 44(3): 109-132. https://doi.org/10.1590/S1806-92902015000300005
Teti N, Hernaman I, Ayuningsih B, Ramdani D, Siswoyo (2018). Pengaruh imbangan protein dan energi terhadap kecernaan nutrien ransum domba garut betina. J. Ilm. Tek. Pet., 6(2): 97–101. https://doi.org/10.23960/jipt.v6i1.p97-100
Tillman AD, Hartadi H, reksodiprojo S, Prawirokusumo, Labdosoekojo S (2005). Ilmu Makanan Ternak Dasar. Gajah Mada University Press. Jakarta.
Usman Y, Eka MS, Nurul F (2013). Evaluasi pertambahan bobot badan sapi aceh jantan yang diberi imbangan antara hijauan dan konsentrat di balai pembibitan ternak unggul indrapuri. Agripet, 13(2): 47-52. https://doi.org/10.17969/agripet.v13i2.819
Van Soest PJ, Robertson, Lewis BA (1991). Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74(10): 3583–3597. https://doi.org/10.3168/jds.S0022-0302(91)78551-2
Wiltbank M, Lopez H, Roberto S, sangriyavong S, Gumen A (2006). Changes in reproductive physiology of lactating dairy cows due to elevated steroid metabolism. Theriogenology, 65(1): 17-29. https://doi.org/10.1016/j.theriogenology.2005.10.003
Wiryawan KG, Saefudin A, Fuah AM, Priyanto R, Khotijah L, Suharti S (2017). Fermentation characteristics and nitrogen retention of madura cattle fed complete rations containing soybean pod and by-products. Med. Pet., 40(1): 28–34. https://doi.org/10.5398/medpet.2017.40.1.28
Yendraliza (2013). The influence of nutrition in the animal reproduction management (A literature review). Kutu D Mhanah, 16: 20-26.
Yetmaneli, Purwanto BP, Priyanto R, Manalu W, Pazla R (2023). Effect of temperaturehumidity index on thermoregulation responses of Pesisir cattle in different altitudes. Int. J. Vet. Sci., 12(6): 879-886.
Zain M, Despal, Tanuwiria UH, Pazla R, Putri EM, Amanah U (2023). Evaluation of legumes, roughages, and concentrates based on chemical composition, rumen degradable and undegradable proteins by in vitro method. Int. J. Vet. Sci., 12(4): 528-538. https://doi.org/10.47278/journal.ijvs/2022.218
Zain M, Putri EM, Ningrat RWS, Erpomen, Makmur M (2020). Effects of supplementing Gliricidia sepium on ration based ammoniated rice straw in ruminant feed to decrease methane gas production and to improve nutrient digestibility (in-vitro). Int. J. Adv. Sci. Eng. Inf. Tec., 10(2): 724–729. https://doi.org/10.18517/ijaseit.10.2.11242
Zhou XQ, Zhang YD, Zhao M, Zhang T, Zhu D, Bu DP, Wang JQ (2015). Effect of dietary energy source and level on nutrient digestibility, rumen microbial protein synthesis, and milk performance in lactating dairy cows. J. Dairy Sci., 98(10): 7209–7217. https://doi.org/10.3168/jds.2015-9312
To share on other social networks, click on any share button. What are these?


