

Effects of Flight Disturbance on Bird Communities at Airports: Predatory Birds Rise to the Challenge





Effects of Flight Disturbance on Bird Communities at Airports: Predatory Birds Rise to the Challenge
Jiaojiao Wang, Wei Liu, Qingqian Lin and Jianhua Hou*
College of Life Science, Hebei University, Baoding 071000, China
ABSTRACT
Airports are special ecological environments. To study the effects of aircraft flight noise disturbance on airport bird communities, the composition and diversity of airport bird communities on flight and non-flight days were investigated using a line transect sampling method at the eastern airport of Hebei Province from June to August, 2019. In total, 36 species belonging to 11 orders and 22 families of birds were recorded. Disturbances such as noise from aircraft flights affected the number of species and the individual density and diversity of birds. However, different areas were affected to different extents, and three areas on the same side were the most affected. Additionally, we found that the numbers of 14 bird species increased on flight days compared to those on non-flight days. Three bird species, upland buzzard Buteo hemilasius, eastern buzzard Buteo japonicus, and black-crowned night heron Nycticorax nycticorax, were only observed on flight days. On flight days, the increase in the number of some raptors may be for hunting. Therefore, to prevent bird collisions, measures should be taken according to specific areas; in addition, bird repellent measures should be applied for bird species that increase during flight days.
Article Information
Received 13 September 2022
Revised 28 September 2022
Accepted 18 October 2022
Available online10 January 2023
(early access)
Published 06 February 2024
Authors’ Contribution
JH designed the study. QL performed the field experiments. JW and WL carried out statistical analyses and wrote the draft manuscript. All authors approved the final submission.
Key words
Flight disturbance, Noise, Bird collisions, Airports, Bird communities
DOI: https://dx.doi.org/10.17582/journal.pjz/20220913080930
* Corresponding author: hbdxnl@163.com
0030-9923/2024/0002-0845 $ 9.00/0
Copyright 2024 by the authors. Licensee Zoological Society of Pakistan.
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Introduction
Anthropogenic noise is becoming more prevalent because of rapid economic development. For many species, noise is an evolutionary source of acoustic disturbance and is potentially an important force affecting the ecology and evolution of many animals (Slabbekoorn and Ripmeester, 2008). Studies have shown that noise affects animal mate choice, song (Derryberry et al., 2020; Zhao et al., 2021), anti-predation (Chan et al., 2010), and other behaviors. For example, the minimum frequency of song of white-crowned sparrow (Zonotrichia leucophrys) increased with increased noise because of increased traffic, and decreased during the COVID-19 shutdown period, when urban noise was decreased (Derryberry et al., 2020). Further, noise levels can affect species diversity, richness, and community structure (Francis et al., 2009; Proppe et al., 2013; Perillo et al., 2017; Carral-Murrieta et al., 2020). However, some studies have found that noise has little or no effect on bird diversity (Yang et al., 2020). Currently, the effects of anthropogenic noise on birds have received increasing attention but are not well-studied.
Urbanization can have a great negative effect on birds, but it can also have some positive effects (Morelli et al., 2014). Airports are special ecological environments, and with rapid growth of the aviation industry, the number of routes and flights is increasing; this raises the issue of noise generated during the landing and take-off cycle (LTO) and by various ground operation equipment during the LTO of aircrafts (Huang, 2020). In addition, collisions between birds and aircraft during take-off, flight, and landing can lead to flight safety accidents or incidents (bird strikes). As bird strikes cause significant damage to aircraft and people’s lives, the International Aviation Federation has upgraded the airport bird strike hazard to an “A” category aviation disaster (Liu, 2011). Current research on airports focuses on aspects such as the spatial and temporal distribution of birds (Hou et al., 2001; Xu et al., 2014), seasonal patterns of bird species (Ma et al., 2015; Chen and Deng, 2020; Liu et al., 2020), bird diversity analysis (Cheng et al., 2020; Li et al., 2020), and bird strike risk assessment (Wang et al., 2010; Zhang et al., 2019). The special environment of airports with large areas of grass and other vegetation attracts birds to perch and feed; however, current research on bird diversity has focused on the effects of urbanization noise or building products of urbanization (e.g. roads) on birds (Morelli et al., 2014; Perillo et al., 2017; Carral-Murrieta et al., 2020; Yang et al., 2020; Wilson et al., 2021). The effects airport noise on the community composition and structure of birds have rarely been reported. Therefore, comparing bird communities and composition in airports on flight days vs. non-flight days will provide insights into the effects of airport noise on birds as well as facilitate the design of targeted measures to prevent bird strikes on flight days.
To this end, the bird community composition and diversity on airport flight days and non-flight days were studied using the line transect sampling method at an airport in eastern Hebei province, China, under good weather conditions from June to August, 2019. We hypothesized that the species, number, and diversity of birds would decrease on airport flight days owing to the influence of airport noise, whereas the species and number of birds would increase on non-flight days; however, the magnitude of its impact in different areas may differ.
Materials and Methods
Overview of the study area
The study airport is located in the eastern plain of Hebei Province, at the lower end of the North China Alluvial Plain, with a gentle topography. The airport has a warm, temperate, semi-humid monsoon continental climate. There is much wind and little rain in the spring, with high evaporation, dry air, and drought. The summer is hot and humid, with concentrated rainfall, heavy rain, hail, thunderstorms, high winds, and severe weather. The autumn is mostly sunny, with large temperature changes, rapid cooling, low wind speed, and fresh air. In winter, the weather is relatively cold and dry, precipitation is scarce, with a prevalent northwest wind. The annual sunshine duration ranges from 2600 to 2900 hours, the annual average temperature is 12.5 ℃; the highest temperature is 32.9 ℃ and the lowest is -14.8 ℃. The frost-free period lasts for 180 to 190 days; annual precipitation is 500 to 700 mm, and the average number of frost days is about 10 days per year.
Survey sample area setting
The airport habitat was dominated by lawns. We first selected 8 sample regions from west to east, namely A to H, and named the four sample regions located in the north as A, C, E and G. The four sample areas located in the south are B, D, F and H. At both ends of these eight areas, J and I areas are respectively set, with a total of 10 sample areas. The reasons for dividing the airport into different areas are as follows: different areas may have different bird species and numbers due to the different intensity of bird repellent activities; according to different areas, the airport can take corresponding measures to prevent bird strikes.
Data acquisition
In the summer of 2019 from June to August, under good weather conditions, the 10 sample areas at the airport were surveyed using the line transect sampling method. Overall, six flight days (two flight days randomly selected each month) and four non-flight days (two non-flight days randomly selected in July and August) were selected for the survey; the survey time was from 06:00 to 18:00 each day, and the survey was conducted once every hour. The survey was divided into 2 groups of 1 person each, and was conducted simultaneously, i.e., one group traveled along the center line of areas A-C-E-G-I, and the other group traveled along the center line of areas J-B-D-F-H simultaneously. The length of each sample area was 500 m, and the observation distance was 50 m on one side. The survey was conducted using an SUV with a speed of about 2.5 km/h and equipment including binoculars (8 × 42), GPS handheld locators, and digital cameras (100–400 mm) to record the species and numbers of birds observed in each sample line. To avoid double counting, birds with the same direction of travel were not counted.
Statistical methods
(1) Bird species dominance: expressed as a percentage (Pi) of the number of individuals to the total number of individuals of different species.
(2) Density of individual birds: D = N/2LS
Where D is the density of individual birds; N is the number of birds recorded; L is the length of the sample area; and S is the one-sided observation distance (Gao, 1993).
(3) Species diversity was calculated using the Shannon-Wiener index (Shannon, 1949).
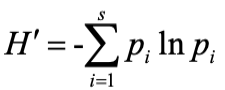
Where H’ is the diversity index, S is the total number of bird species, and Pi is the number of individuals of the ith species as a proportion of the total number of individuals.
(4) Uniformity was calculated using the Pielou index (Pielou, 1975).
J = H’ / ln s
Where H’ is the Shannon-Weiner index and S is the number of species.
(5) Population ranking: Calculated using the percentage of the population, i.e., the number of a certain bird species/total number of all birds × 100%, defining species with more than 10% as the dominant species, species with 1–10% as common species, and species with less than 1% as rare species.
(6) Feeding groups division (Teresa, 1999): (1) grain-eating birds: Birds that feed mainly on plant seeds or crop seeds, such as corn and wheat; (2) insectivorous birds: birds that feed mainly on adult insects, larvae and worms; (3) carnivorous birds: birds that feed mainly on small vertebrates such as mice and small birds; (4) omnivorous birds: birds with a mixed diet, mainly feeding on animal and plant foods.
(7) Data analysis: Generalized linear models (GLMs) were used to analyze the effects of aircraft flight noise on the number of bird species, individual density, species diversity, and uniformity at the airport as targets, and whether the plane flew (yes or not), the area, and the interaction of the two as fixed effects. We also used GLMs to investigate the effects of aircraft flight on bird densities and bird species numbers at different times of the day, and because the aircraft took off at different times of the day during flight days, three times of the six flight days surveyed (i.e., 8 am–9am, 9 am–10 am, and 10 am–11 am) were selected for comparison with non-flight days, considering the bird species numbers and individual densities as responses. Whether the plane flew, the time period, and the interaction of the two were inserted into the model as fixed effects.
Results
Bird species composition
The survey recorded 36 species of birds belonging to 11 orders and 22 families at the airport. Of these, 11 families and 16 species of birds belonged to the order Passeriformes and 11 families and 20 species of birds were non-Passeriformes (Supplementary Table SI).
Based on the population numerical rank, the dominant species were eurasian tree sparrow (Passer montanus) and barn swallow (Hirundo rustica); five common species included common magpie (Pica pica), rock dove (Columba livia domestica), white-cheeked starling (Spodiopsar cineraceus), Richard’s pipit (Anthus richardi), and pintail snipe (Gallinago stenura).
Based on the feeding group, there were 7 species of omnivorous birds, accounting for 19% of all bird species at the airport; further, there were 16 species of insectivorous birds, accounting for 45% of all bird species; 5 species of grain-eating birds, accounting for 14% of bird species; and 8 species of carnivorous birds, accounting for 22% of the number of bird species (Fig. 1, Supplementary Table SI).
Effects of flight on birds at different times of the day
The GLM results showed no significant effects regarding whether the aircraft flew (x2 = 1.519, df = 1, P = 0.218), the different time periods (x2 = 1.379, df = 2, P = 0.502), or the interaction between whether the aircraft flew and time period (x2 = 1.716, df = 2, P = 0.424) on bird species numbers (Table I).
Table I. Influence of interference factors (whether the plane flew, area and the interaction between the two) on the number of bird species, individual density, species diversity and evenness at the airport.
|
Source |
X2 |
df |
P |
|
Model for the number of bird species |
|||
|
Whether the plane flew |
31.736 |
1 |
< 0.001 |
|
Area |
49.297 |
9 |
< 0.001 |
|
Whether the plane flew x area |
12.219 |
9 |
0.201 |
|
Model for individual density of birds |
|||
|
Whether the plane flew |
52.839 |
1 |
< 0.001 |
|
Area |
64.092 |
9 |
< 0.001 |
|
Whether the plane flew x area |
44.271 |
9 |
< 0.001 |
|
Model for species diversity |
|||
|
Whether the plane flew |
0.628 |
1 |
0.428 |
|
Area |
7.678 |
9 |
0.567 |
|
Whether the plane flew x area |
20.619 |
9 |
0.014 |
|
Model for uniformity |
|||
|
Whether the plane flew |
6.430 |
1 |
0.011 |
|
Area |
8.229 |
9 |
0.511 |
|
Whether the plane flew x area |
16.646 |
9 |
0.055 |
There was a significant effect of whether the aircraft flew or not on individual density (x2 = 9.775, df = 1, P = 0.002; Table I), with birds on non-flight days having significantly higher individual densities than those on flight days (Fig. 2); however, neither the time period (x2 = 0.535, df = 2, P = 0.765; Table I) nor the interaction between whether the aircraft flew and the time period (x2 = 0.650, df = 2, P = 0.722; Table I) had any significant effect on individual bird density.
Effects on birds in different regions
Effect of aircraft flight on the number of bird species
Whether the aircraft flew or not had a significant effect on the number of bird species (x2 = 31.736, df = 1, P < 0.001; Table I); the number of bird species was significantly higher on non-flight days (18.75 ± 4.79) than that on flight days (12.67 ± 3.08); the number of bird species also differed significantly between the different areas (x2 = 49.297, df = 9, P < 0.001; Table I), with areas I and J having the highest number of bird species whereas areas C, E, and G on the same side had the lowest number of bird species. Pairwise comparisons showed that all differences were attributed to a significantly higher number of bird species in area J than that in all other areas (all P < 0.05) and to the differences between the same-side partitions, i.e., areas C, E, and G, and the other partitions (all P < 0.05), except for the same side area A, which had significantly more bird species than those in area G (P = 0.010). However, the interaction of whether the aircraft flew or not with the partitions did not significantly affect the number of bird species (x2 = 12.219, df = 9, P = 0.201; Table I).
Effect on individual bird densities
There was a significant effect of whether the aircraft flew on the density of individual birds (x2 = 52.839, df = 1, P < 0.001; Table I), which was significantly lower when the aircraft flew (73.39 ± 39.97) than that when it did not fly (366.20 ± 310.09); there was a significant difference between areas in the density of individual birds (x2 = 64.092, df = 9, P < 0.001; Table I), and the interaction between whether the aircraft flew and area partition also had a significant effect (x2 = 44.271, df = 9, P < 0.001; Table I). The density of individual birds when the aircraft was flying and not flying followed roughly the same trend across partitions, with the lowest density of individual birds in areas C, E, and G on the same side. Paired results showed that, in terms of differences between areas when the aircraft was flying or not flying, there were no significant differences in individual densities between areas when the aircraft was flying (all P > 0.05); however, when the aircraft was not flying, differences between areas were mainly attributed to significantly higher individual densities in area J than those in all other areas (all P > 0.05), and significantly lower individual densities in areas C, E, and G on the same side than those in the other areas (all P > 0.05). Further, for each area, individual bird densities were significantly higher in areas D, F, H, I and J when the aircraft were not flying than those when aircraft were flying (all P < 0.05).
Impact on diversity
For bird diversity, only the interaction of whether or not the aircraft flew with the area partition was significant (x2 = 20.619, df = 9, P < 0.014; Table I). Pairwise comparisons showed that for each area, only two areas, E (P = 0.024) and G (P = 0.012), had a significantly higher species diversity when aircrafts were not flying than that when they were flying.
In terms of differences in species diversity between areas with and without aircraft flight, no significant differences in diversity were observed among areas when aircrafts were not in flight (all P > 0.05). When aircrafts were in flight, all variability was caused by significantly lower species diversity in areas A, C, E, and G on the same side than that in the other areas (all P < 0.05), with the exception of area B, where species diversity was significantly higher than in that area D (P = 0.043) and area C, where species diversity was significantly higher than that in area G (P = 0.039).
Effect on uniformity
When analyzing the effect of whether or not aircraft flew on the uniformity of each area, a significant difference was found only for whether or not the aircraft flew on the uniformity of the birds (x2 = 6.430, df = 1, P = 0.011; Table I), which was significantly higher when flying (0.53 ± 0.08) than when not flying (0.49 ± 0.02).
Effects of aircraft flight on different bird species
Bird densities that increased on flight days compared to those on non-flight days included the hen harrier, upland buzzard (Buteo hemilasius), northern lapwing (Vanellus vanellus), amur falcon, common kestrel (Falco tinnunculus), eurasian collared dove (Streptopelia decaocto), white-cheeked starling, rock dove, eastern buzzard (Buteo japonicus), Richard’s pipit, black-crowned night heron (Nycticorax nycticorax), Chinese grey shrike (Lanius sphenocercus), eurasian skylark (Alauda arvensis), and spotted dove (Streptopelia chinensis). Birds that occurred only on flight days included upland buzzard, eastern buzzard, and black-crowned night herons, and birds that occurred only on non-flight days included Swinhoe’s snipe (Gallinago megala), black drongos (Dicrurus macrocercus), gray wagtail (Motacilla cinerea), grey-headed woodpecker (Picus canus), red-rumped swallow (Cecropis daurica), Indian cuckoo (Cuculus micropterus), long-billed plover (Charadrius placidus), and little owl (Athene noctua).
Discussion
Our airport summer bird survey recorded a total of 36 species in 11 orders and 22 families of birds, including 11 families and 16 species of passerine birds and 11 families and 20 species of non-passerine birds. Disturbances such as noise from aircraft flight affected the number of species, individual density, species diversity, and uniformity of birds; with the three areas (C, E and G) on the same side being the most affected; this may be related to aircraft flights and their bird repellent activities. In addition, certain bird species such as upland buzzard, eastern buzzard, and common kestrel were increased in numbers on flight days.
Noise has a significant impact on animal behavior (Derryberry et al., 2020; Zhao et al., 2021) but the effects of airport noise on bird communities and diversity have not been reported. In this study, we recorded the species and numbers of birds at a selected airport on flight days and non-flight days, and found that passerines were high in numbers and dominated the bird community at the airport, consistent with the results of previous studies on the bird community composition at airports in Hebei Province (Ma et al., 2015; Wu et al., 2019). As hypothesized, we found that the number of bird species declined with airport noise disturbance (i.e., flight days) and that the overall number of bird individuals decreased significantly, mainly among eurasian tree sparrow, barn swallow, red-rumped swallow, light-vented bulbul (Pycnonotus sinensis), and azure-winged magpie (Cyanopica cyanus) on flight days, indicating that these small and medium-sized birds may have an active tendency to avoid airport noise. Our results are consistent with studies on the effects of road noise on animal populations (Fahrig and Rytwinski, 2009), indicating that noise exhibits mainly negative or minimal effects on animals. Wang et al. (2011) reported that the population density of all woodland birds started to decline when noise levels reached 42 dB on average. Noise from aircraft flights can reach over 100 dB (Zhang, 2018), and such strong noise may lead to intolerance in some bird species and decrease their population. In contrast, some bird populations did not decrease even in the presence of airport noise, possibly because they had adapted to this environment, i.e., as anthropogenic noise becomes more common, the intensity of animal responses to them may decrease because of habituation (Thompson and Spencer, 1966).
Interestingly, some bird species increased in numbers in the presence of airport noise, including amur falcon, common kestrel, hen harrier, eastern buzzard, upland buzzard, and Chinese grey shrike, while eastern buzzard and upland buzzard only occurred in the presence of airport noise. Some of these birds are carnivorous and may emerge in search of prey when airport noise is present; consistent with the results of this study, other studies have found that animals are attracted to and benefit from noisy, disturbed areas, for example, in search of food (Berger, 2007; Fahrig and Rytwinski, 2009; Morelli et al., 2014). Here, there are three interpretations for the increase in raptor numbers in the presence of airport noise. First, some prey may engage in herding behavior in the absence of noise; the Group Effect Hypothesis (GEH) suggests that each member of a group will use other individuals as a buffer to reduce the probability of being preyed on (Siegfried and Underhill, 1975; Goodale and Ruxton, 2010). In the presence of airport noise, some birds may react and escape in fear (Weisenberger et al., 1996; Maier et al., 1998; Quinn et al., 2006; Rabin et al., 2006; Nowacek et al., 2007), causing such prey to expose itself, increasing the chances of their predation. Some studies have also constructed models and shown that populations are unable to engage in cluster behavior when noise is high (Chen and Liu, 2021; Zhang et al., 2021). Second, noise masks the sound of predators (acoustic masking), allowing them to exploit the situation. Third, noise may effectively distract prey, thus preventing them from responding to approaching predators (distracted prey hypothesis) (Chan et al., 2010). The increase in predatory birds such as raptors found in this study may be similar to our previous study on playback alarms (Wang et al., 2020), which indicated that playback or noise exposes prey targets, which may attract predators. Further, rock dove and dove tend to perch or nest in villages and woodlands surrounding airports, and the increase in the number of their individuals on flight days may be related to them being frightened and entering the airport.
Of the 10 studied sample areas at the airport, three areas (C, E and G) were the most affected by disturbances such as airport noise compared to other areas, which could significantly reduce the number of bird species and individuals in these areas, as this area was affected by aircraft landings and takeoffs as well as taxiing or other bird repelling activities. It is thus clear that airport noise affects birds differently in each area of the airport. Shen (1983) studied the hearing of animals and showed that birds can easily detect vibrations caused by noise inside animals or underground, therefore the noise and vibrations generated by aircraft flight are highly disturbing to birds. The area located on the south side and I and J at both ends are less disturbed by aircraft take-off and landing, therefore the species and number of birds in these areas are relatively less affected. However, the birds in these areas also pose a notable impact on the occurrence of bird strikes. Further, whether the aircrafts are in flight or not, the relatively low number of bird species and numbers in areas C, E, and G, may be influenced by aircraft noise disturbances as well as be related to the surrounding environment. It has recently been shown that noise can affect the reproductive success of fish (Nedelec et al., 2022), and that further research is needed to determine whether noise near airports can affect the breeding of birds. In addition, noise may also affect the frequency of bird calls (Derryberry et al., 2020), which also needs to be investigated further.
Conclusion
Our results indicate that on airport flight days, the overall number of bird species, individual density, and species diversity is relatively low, which may be related to disturbances such as noise from aircraft flight; three areas on the same side are the most affected, and is related to aircraft flight and bird repellent activities. However, other areas should also receive attention to prevent and control the occurrence of bird strikes. Further, the numbers of certain bird species such as upland buzzard, eastern buzzard, and common kestrel increased on flight days; therefore, subsequent studies are needed to focus on the activity patterns of these species on flight days and propose the targeted measures to prevent bird strikes. Moreover, the breeding success and bird calls around the airport need to be further studied in terms of how they are affected by airport noise.
Acknowledgements
We thank the airport in eastern Hebei province for support and permission to carry out this study. We also thank the anonymous reviewers whose feedback improved the quality of our manuscript.
Funding
This work was funded by the Animal Ecology Innovation Team Project of Hebei University (2021-22) and Life Science and Green Development Discipline Group Project of Hebei University (2021-2022).
IRB approval and ethical statement
This experiment only observed the birds at the airport, therefore, no ethical approval and Institutional review was required.
There is supplementary material associated with this article. Access the material online at: https://dx.doi.org/10.17582/journal.pjz/20220913080930
Statement of conflict of interest
The authors have declared no conflict of interest.
References
Berger, J., 2007. Fear, human shields and the redistribution of prey and predators in protected areas. Biol. Lett., 3: 620-623. https://doi.org/10.1098/rsbl.2007.0415
Carral-Murrieta, C.O., García-Arroyo, M., Marín-Gómez, O.H., Sosa-López, J.R., and MacGregor-Fors, L., 2020. Noisy environments: untangling the role of anthropogenic noise on bird species richness in a Neotropical city. Avian Res., 11: 32. https://doi.org/10.1186/s40657-020-00218-5
Chan, A.A.Y.H., Giraldo-Perez, P., Smith, S., and Blumstein, D.T., 2010. Anthropogenic noise affects risk assessment and attention: The distracted prey hypothesis. Biol. Lett., 6: 458-461. https://doi.org/10.1098/rsbl.2009.1081
Chen, B., and Deng, B., 2020. Bird biodiversity and seasonal changes at nanchang Changbei international airport. Chin. J. Wildl., 41: 359-368. https://doi.org/10.19711/j.cnki.issn2310-1490.2020.02.015
Chen, M., and Liu, Y., 2021. Flocking dynamics of a coupled system in noisy environments. Stochastics Dyn., 21. https://doi.org/10.1142/S0219493721500568
Cheng, L., Wang, X., Xu, Y., Cai, X., Xue, W., and Wan, X., 2020. Characteristics of avian diversity and birdstrike prevention and control at Taiyuan Wusu international airport. Chin. J. Ecol., 39: 2014-2023. https://doi.org/10.13292/j.1000-4890.202006.017
Derryberry, E.P., Phillips, J.N., Derryberry, G.E., Blum, M.J., and Luther, D., 2020. Singing in a silent spring: Birds respond to a half-century soundscape reversion during the COVID-19 shutdown. Science, 370: 575-579. https://doi.org/10.1126/science.abd5777
Fahrig, L., and Rytwinski, T., 2009. Effects of roads on animal abundance: An empirical review and synthesis. Ecol. Soc., 14: 21. https://doi.org/10.5751/ES-02815-140121
Francis, C.D., Ortega, C.P., and Cruz, A., 2009. Noise pollution changes avian communities and species interactions. Curr. Biol., 19: 1415-1419. https://doi.org/10.1016/j.cub.2009.06.052
Gao, W., 1993. Avian Ecology. Northeast Normal University Press, Changchun.
Goodale, E., and Ruxton, G.D., 2010. Antipredator benefits from heterospecifics. In: Encyclopedia of Animal Behavior (eds. M.D. Breed and J. Moore). Academic Press, Oxford. pp. 94-99. https://doi.org/10.1016/B978-0-08-045337-8.00333-8
Hou, J., Wu, M., Hu, Y., Zhang, X., and Hu, Z., 2001. Study on the bird community structure in the forest-steppe transition zone. Acta Zool. Sin., 47: 148–156.
Huang, X., 2020. Impacts of airport noise pollution on the surrounding ecosystem and planning. Res. Chin. Constr., 6: 90-91.
Li, W., Hong, L., Wang, G., Zeng, W., Wang, Y., Li, B., Li, D. and Luo, X.2020. Avain diversity and bird strike risk evaluation in Ninglang Luguhu Airport, Yunnan. J. Southw. For. Univ., 40: 96-102. https://doi.org/ 10.11929/j.swfu.201903150
Liu, A., Zheng, Y., Xu, Q., and Zhang, M., 2020. Spring bird community and bird strike prevention at Qiqihar airport, Heilongjiang, China. Chin. J. Wildl., 41: 349-358. https://doi.org/ 10.19711/j.cnki.issn2310-1490.2020.02.014
Liu, Z., 2011. The causes of bird strike and the preventive strategies. Chian Civil Aviation., 130: 62-64.
Ma, L., Chen, C., and Hou, J., 2015. Bird community distribution and activity rhythm on airport in the east of Hebei in autumn. J. Tianjin Norm. Univ. (Natl. Sci. Ed.). 35: 155-159. https://doi.org/ 10.3969/j.issn.1671-1114.2015.03.039
Maier, J.A.K., Murphy, S.M., White, R.G., and Smith, M.D., 1998. Responses of caribou to overflights by low-altitude jet aircraft. J. Wildl. Manag., 62: 752-766. https://doi.org/10.2307/3802352
Morelli, F., Beim, M., Jerzak, L., Jones, D., and Tryjanowski, P., 2014. Can roads, railways and related structures have positive effects on birds? A review. Transp. Res. D Transp. Environ., 30: 21-31. https://doi.org/10.1016/j.trd.2014.05.006
Nedelec, S.L., Radford, A.N., Gatenby, P., Davidson, I.K., Velasquez Jimenez, L., Travis, M., Chapman, K.E., McCloskey, K.P., Lamont, T.A.C., Illing, B., McCormick, M.I. and Simpson, S.D., 2022. Limiting motorboat noise on coral reefs boosts fish reproductive success. Nat. Commun., 13: 2822. https://doi.org/10.1038/s41467-022-30332-5
Nowacek, D.P., Thorne, L.H., Johnston, D.W., and Tyack, P.L., 2007. Responses of cetaceans to anthropogenic noise. Mammal. Rev., 37: 81-115. https://doi.org/10.1111/j.1365-2907.2007.00104.x
Perillo, A., Mazzoni, L.G., Passos, L.F., Goulart, V.D.L.R., Duca, C., and Young, R.J., 2017. Anthropogenic noise reduces bird species richness and diversity in urban parks. Int. J. Avian Sci., 159: 638-646. https://doi.org/10.1111/ibi.12481
Pielou, E.C., 1975. Ecological diversity. John Wiley and Sons Inc., New York.
Proppe, D.S., Sturdy, C.B., and St Clair, C.C., 2013. Anthropogenic noise decreases urban songbird diversity and may contribute to homogenization. Glob. Change Biol., 19: 1075-1084. https://doi.org/10.1111/gcb.12098
Quinn, J.L., Whittingham, M.J., Butler, S.J., and Cresswell, W., 2006. Noise, predation risk compensation and vigilance in the chaffinch Fringilla coelebs. J. Avian Biol., 37: 601–608. https://doi.org/10.1111/j.2006.0908-8857.03781.x
Rabin, L.A., Coss, R.G., and Owings, D.H., 2006. The effects of wind turbines on antipredator behavior in California ground squirrels (Spermophilus beecheyi). Biol. Conserv., 131: 410-420. https://doi.org/10.1016/j.biocon.2006.02.016
Shannon, C.E., 1949. The mathematical theory of communication. University of Illinosi Press, Urbana.
Shen, J., 1983. A behavioral study of vibrational sensitivity in the pigeon (Columba livia). J. comp. Physiol., 152: 251-255. https://doi.org/10.1007/BF00611189
Siegfried, W.R., and Underhill, L.G., 1975. Flocking as an anti-predator strategy in doves. Anim. Behav., 23: 504-508. https://doi.org/10.1016/0003-3472(75)90126-8
Slabbekoorn, H., and Ripmeester, E.A., 2008. Birdsong and anthropogenic noise: Implications and applications for conservation. Mol. Ecol., 17: 72-83. https://doi.org/10.1111/j.1365-294X.2007.03487.x
Teresa, F., 1999. Relationships of avian cecal lengths to food habits, taxonomic position, and intestinal lengths. Condor, 101: 622-634. https://doi.org/10.2307/1370192
Thompson, R.F., and Spencer, W.A., 1966. Habituation: A model phenomenon for the study of neuronal substrates of behavior. Psychol. Rev., 73: 16-43. https://doi.org/10.1037/h0022681
Wang, J., Ma, L., Liang, W., and Yang, C., 2020. Responses of cuckoo hosts to alarm signals of different nest intruders in non-nesting areas. Zool. Res., 41: 345-350. https://doi.org/10.24272/j.issn.2095-8137.2020.030
Wang, L., Zhou, L., Wan, Z., Zhang, B., Xu, R., Zhang, Y. and Xue, W., 2010. Avian diversity and bird strike risks on Luogang Airport. Chin. J. Wildl., 31: 127-130. https://doi.org/ 10.19711/j.cnki .issn2310 -1490.2010.03.004
Wang, Y., Li, Q., Guan, L., Fang, R., and Jiang, R., 2011. Effect of traffic noise around napahai wetland highway on birds. Chin. J. Zool., 46: 65–72. https://doi.org/ 10.13859/j.cjz.2011.06.013
Weisenberger, M.E., Krausman, P.R., Wallace, M.C., De Young, D.W., and Maughan, O.E., 1996. Effects of simulated jet aircraft noise on heart rate and behavior of desert ungulates. J. Wildl. Manag., 60: 52-61. https://doi.org/10.2307/3802039
Wilson, A.A., Ditmer, M.A., Barber, J.R., Carter, N.H., Miller, E.T., Tyrrell, L.P. and Francis, C.D., 2021. Artificial night light and anthropogenic noise interact to influence bird abundance over a continental scale. Glob. Change Biol., 27: 3987-4004. https://doi.org/10.1111/gcb.15663
Wu, T., Wang, Y., Ren, H., Gao, L., and Hou, J., 2019. Commincity composition and dynamic of bird in the airport. J. Hebei Univ. (Nat. Sci. Ed.). 39: 175–182. https://doi.org/10.3969/j.issn.1000-1565.2019.02.010
Xu, Q., Zhang, X., Song, Y., Wang, C., and Chang, L., 2014. The Vertical Distribution of Birds in Flight at Qiqihar Airport Airspace in Spring and Autumu. Chin. J. Wildl., 35: 194-199. https://doi.org/ 10.19711/j.cnki .issn2310 -1490.2014.02.016
Yang, X., Tan, X., Chen, C., and Wang, Y., 2020. The influence of urban park characteristics on bird diversity in Nanjing, China. Avian Res., 11. https://doi.org/10.1186/s40657-020-00234-5
Zhang, Q., He, M., Zhao, X., He, X., Dou, L., and Ran, J., 2019. A study on the relationship between bird strike risk and bird community in Chengdu Shuangliu international Airport. Sichuan J. Zool., 38: 576-586. https://doi.org/10.11984/j. issn. 1000-7083. 20180179
Zhang, Y., 2018. Airport Noise assessment and mitigation strategies. Netland, 1: 170-214. https://doi.org/ 10.3969/j.issn.1672-528X.2018.01.157
Zhang, Y., Zhou, R., and Chen, X., 2021. Dynamic behavior of class of flocking models disturbed by white noise. J. Jilin Univ. (Nat. Sci. Ed.). 59: 1345-1350. https://doi.org/10.13413/j.cnki.jdxblxb.2021135
Zhao, L., Wang, T., Guo, R., Zhai, X., Zhou, L., Cui, J. and Wang, J., 2021. Differential effect of aircraft noise on the spectral-temporal acoustic characteristics of frog species. Anim. Behav., 182: 9-18. https://doi.org/10.1016/j.anbehav.2021.09.014
To share on other social networks, click on any share button. What are these?



