
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 1 (3S): 25 – 29Special Issue–3 (Epidemiology and Animal Disease Investigations)
Hydropericardium Syndrome (HPS) Virus: Immunofluorescence Studies on Aspects of Pathogenesis in Chickens
Rajesh Kumar1*, Rajesh Chandra2, Vipan Kumar3, Prakash Bhatt4, Santosh Kumar Shukla5, Kuldeep Dhama6
- Department of Veterinary Microbiology, College of Veterinary and Animal Sciences, G.B. Pant University of Agriculture & Technology, Pantnagar – 263 145, Uttarakhand
- Department of Veterinary Microbiology, College of Veterinary & Animal Sciences, Central Agricultural University, Selseih, Aizawal
- Department of Veterinary Microbiology, College of Veterinary & Animal Sciences, Khalsa College of Veterinary and Animal Sciences, Amritsar, Punjab
- Veterinary Clinics, College of Veterinary and Animal Sciences, G.B. Pant University of Agriculture & Technology, Pantnagar – 263 145, Uttarakhand
- Veterinary Medicine, College of Veterinary and Animal Sciences, G.B. Pant University of Agriculture & Technology, Pantnagar – 263 145, Uttarakhand
- Division of Pathology, Indian Veterinary Research Institute, Izatnagar
*Corresponding author:rajeshvet@rediffmail.com
ARTICLE CITATION:
Kumar R, Chandra R, Kumar V, Bhatt P, Shukla SK and Dhama K (2013). Hydropericardium syndrome (HPS) virus: immunofluorescence studies on aspects of pathogenesis in chickens. Adv. Anim. Vet. Sci. 1 (3S): 25 – 29.
Received: 2013–11–07, Revised: 2013–12–30, Accepted: 2013–12–31
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/198/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Three week old chickens– not previously exposed to hydropericardium syndrome virus (HPSV), were HPSV inoculated by oral and intramscular routes. Chicks were necropsied on day 1, 2, 3, 5 and 10 following inoculation, and specimens from various tissues were collected for examination. There were no apparent clinical signs and mortality in the chicks throughout the experimental period. The distribution and localization viral antigens in various tissues and peripheral blood smears were studied by indirect immunofluorescent technique. Antibody titers during the course of infection were also determined by enzyme linked immunosorbent assay (ELISA). Virus spread to most of organs within 3–5 days post infection and a direct relationship between appearance of antibodies and recovery from infection was recorded.
INTRODUCTION
Fowl adenoviruses (FAdV) cause many disease conditions like inclusion body hepatitis (IBH), hydropericardium syndrome (HPS) respiratory disease, tenosynovitis, impaired growth, reduced egg production, aplastic anemia, atrophy of bursa and thymus enteritis and conjunctivitis in poultry and other birds (Meulemans et al., 2001; Kumar et al., 2003a, 2003b; McFerran and Adair, 2003; Kumar et al., 2010). All eleven serotypes of FAdV have been isolated from natural infections of poultry and other birds and strains of FAdV4 cause hyderopericardium syndrome in chicken (Choi et al., 2012; Asthana et al., 2013).
Hydropericardium syndrome, a disease primarily of broiler birds, has been a severe hazard to poultry industry since it was diagnosed in 1987. During field outbreaks, almost all exposed susceptible broilers may be affected and the mortality may range from 20% to 80% (Akhtar et al., 1992; Asthana et al., 2013). The accumulation of straw/amber coloured jelly–like fluid in the pericardial sac mainly characterizes the disease. Other predominant lesions include enlarged and discoloured liver with foci of haemorrhage and/or necrosis and enlarged kidneys with distended tubules (Abe et al., 1998; Anjum et al., 1989; Calnek and Hitchner, 1969). Histological, electron microscopic and serological studies have shown that causative agent is a Fowl adenovirus serotype 4 (Asthana et al., 2013). In this report, we present the results of a study on aspects of pathogenesis of HPS following oral and intramascular administration of a field isolate (HPS–R) to 3–week–old chickens. We used indirect immunofluorescent technique to detect viral antigens in a range of tissues and ELISA to study humoral immune response during course of infection.
MATERIALS AND METHODS
Virus
An isolate of HPS virus recovered from natural outbreak and propagated in chicken embryo liver cell (CEL) and characterized serologically (Kumar et al., 2003b) was used in the present study.
Experimental Chicks
Day old broiler chicks were procured from a commercial hatchery at Haldwani, Uttarakhand. The chicks were reared on deep litter in an isolation unit until 3–weeks–old and then transferred to battery cages before inoculation. Uninoculated control chicks and infected chicks were kept in separate isolation unit.
Preparation of Hyperimmune Serum
Hyperimmune serum was produced against reference virus by subcutaneous inoculation of 3–week–old chickens with 20% (w/v) liver homogenate extract as described earlier (Ravikumar et al., 1997) for use in the present study. Gamma globulins from hyperimmune serum were purified and incubated with equal volume of a 10% healthy chicken liver cell suspension for 1 hour at room temperature (Saifuddin and Wilks, 1990).
Indirect Fluorescent Antibody Test
The peripheral blood and tissue sections were collected and prepared as described by Fasina and Fabricant (1982). The staining procedure and scoring system were followed as described by earlier workers (Calnek and Hitchner, 1969; Fasina and Fabricant, 1982). In all cases, only relatively homogeneous intranuclear fluorescence was considered positive. Sometimes fluorescent particulate material could be observed in the cytoplasm of cells from both infected and control birds, which was considered a non–specific artifact.
Experimental Design
Two groups of 10 and one control group of 05 chicks were made from 25 birds of three weeks age. All chicks were tested for presence of antibodies against HPSV prior to commencement of experiment. Chicks in group I were inoculated orally with CEL cell culture supernatant (10th passage virus with a titer of log10 6.2 TCID50 /ml) @ 0.75 ml/chick by oral route. Chicks in Group II were inoculated with same virus preparation by intramuscular (i.m.) route. Group III chicks were kept as uninoculated control.
All groups were reared separately under deep litter system during the entire course of experiment. Different organs, viz., liver, kidney, heart, spleen, lung, bursa, intestine and caecum were collected by sacrificing two chicks each from group I & II and one chick from group III on days 1, 2, 3, 5 and 10 days post inoculation. All the organs were kept at –800C until processed for indirect fluorescent antibody test (IFAT). Before necropsy, blood smears were made from each chicken for detection of viral antigens. Blood was also collected on each day for determination of serum antibody titers by ELISA as per the method of Kumar et al. (2003a).
RESULTS
Clinical Signs and Postmortem Lesions
There were no apparent clinical signs in the chicks throughout the experimental period. However, postmortem examination indicated that the liver was enlarged and dark red and showed necrotic foci from 2nd day post infection (DPI) onwards while kidneys were pale and haemorrhagic and changes were visible from 3rd DPI. Congestion of heart was observed on day 2, 3 and 5 DPI and about 2.0 ml of fluid could be aspirated from pericardial sac on day 2 and 5 PI. Enlargement of bursa of Fabricius and spleen and congestion of lungs were observed upto 10th DPI. The spleen also showed necrotic foci on day 5 and 10 PI, however, intestine and caecum did not exhibit any appreciable change on any day PI. On day 10 PI, haemorrhagic foci in breast muscles were recorded in birds infected by i.m. route. In addition, adhesions in the heart were also evident.
Viremia
Viral antigens were not detected in blood smears from chicks inoculated via oral route, however, viral antigens were detected extracellularly from chicks inoculated via i.m. route in less amount on day 1 PI. On day 2 and day 3 PI the blood smears prepared from the chicks infected via both the routes revealed intracellular as well as extracellular presence of viral antigens. However, in comparison to day 2PI, amount of antigen appeared to be less at day 3 PI. On day 5 and 10 PI viral antigens were detected only extracellularly in blood smears prepared from chicks infected via both routes. Table 1 and 2 show the detailed results of the studies on localization of HPS viral antigen in tissues of chickens infected with both the routes.
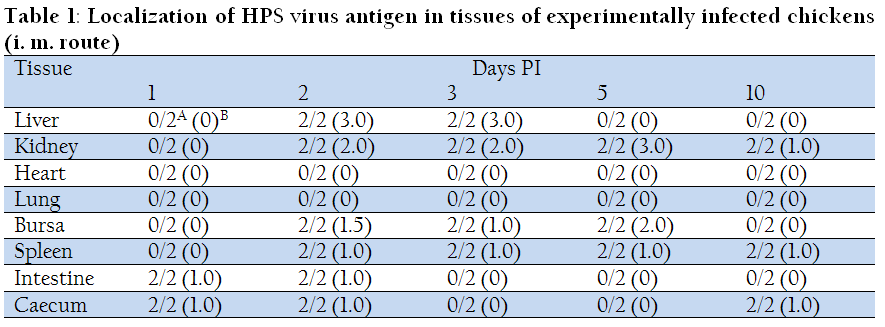
Table 1: Localization of HPS virus antigen in tissues of experimentally infected chickens (i. m. route)
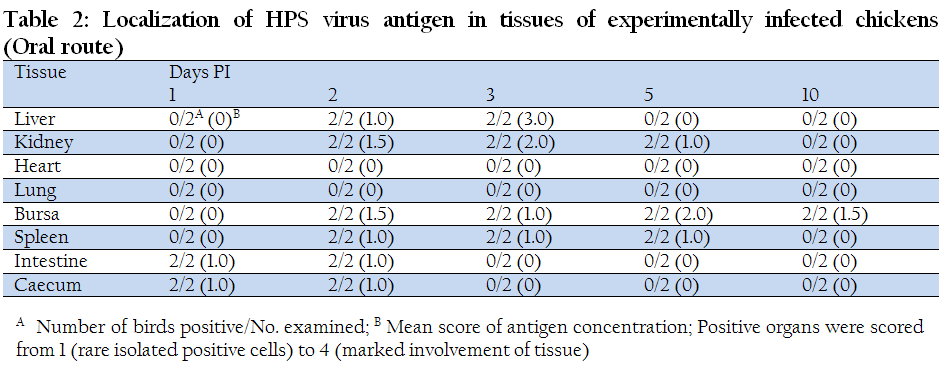
Table 2: Localization of HPS virus antigen in tissues of experimentally infected chickens (Oral route)
Viral Antigen in Tissues and Antibody Response
Of the two chickens sacrificed at one day PI, viral antigens could not be demonstrated in the tissues by IFAT except caecum and intestine from birds infected via oral route. On day 2 PI, the liver, kidney, spleen, intestine, caecum and bursa of Fabricius showed presence of viral antigens. Observations were similar for both the routes of inoculation but, in liver and kidney, involvement of tissue was more in chicks infected via intramuscular route in comparison to oral route. The tissues in other organs showed more or less similar changes, however, lung and heart did not show presence of viral antigens. On day 3 PI, only liver, kidney, bursa of Fabricius and spleen revealed presence of viral antigens. Amount of antigen, as indicated by lesser numbers of fluorescent foci, was less in comparison to day 2 PI. Involvement of tissue was quite similar in both chicks infected via both routes. Viral antigens were not detected in heart, intestine, caecum and lungs. On day 5PI, viral antigens were detected in bursa of Fabricius, kidney and spleen and not detected in liver, lung, caecum and intestine. Involvement of tissue was more in kidney tissue from birds infected via oral route as compared to intramuscular route. On day 10 PI, viral antigens were detected in kidney spleen and caecum and not detected in liver, heart, lung, bursa of Fabricius and intestine from chicks infected by i.m. route, while viral antigens were detected only in the spleen of chicks infected via oral route.
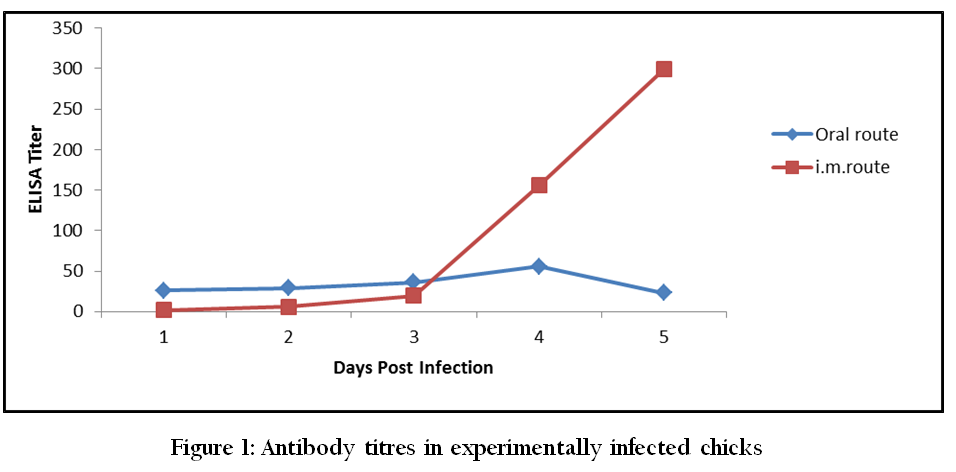
Prior to administration of the virus, serum from all birds was tested negative for antibodies to HPSV. In chicks inoculated via oral route the antibody titres at 1, 2, 3, 5 and 10 days PI were 26.02, 28.96, 36.04 55.84 and 23.29, respectively. ELISA titres recorded in chicks infected via i.m. route at 1, 3, 5 and 10 days PI were 1.48, 19.45, 155.47 and 299.04, respectively (Figure 1).
DISCUSSION
The predominant post–mortem lesions, as described by several investigators (Abdul–Aziz and Hasan, 1995; Asrani et al., 1997; Kumar et al., 2004; Kumar and Chandra, 2009), were also observed in present study. However, dilation of right ventricle seen in ascites syndrome was not noted in the present investigation and viral antigens were not detected in heart tissue. Therefore, it seems logical that cause of hydropericardium in HPS may be different from that of ascites (Abe et al., 1998). Generally in animals other than chickens, chronic disease induces hyfropericardium. However, in hydropericardium syndrome, the period to form hydropericardium is very short. Thus it appears that resultant acute hydremia and hypoproteinemia may cause hydropericardium in chickens. Hydropericardium may also be caused by immediate massive effusion of fluid through capillaries of epicardium into the pericardial sac. Hydropericardium itself can depress cardiac function resulting into muffing of heart sounds and if severe produces a defect in filling and low pressure (Fraser et al., 1991; Nakamura et al., 1999).
In the present study an attempt was made to study the pathogenesis of hydropericardium syndrome virus in broiler chicks of 3 weeks of age by IFAT. Two different routes of inoculation viz., oral and i.m. were used to infect chicks with chicken embryo liver (CEL) cell culture grown virus. IFAT of blood smears suggested that virus was mostly present extracellularly in blood plasma, however, blood cells probably lymphocytes revealed intracellular fluorescence suggesting that some amount of virus was present within these cells. Saiffudin and Wilks (1991) reported that viral antigens were detected only in the plasma fraction of blood from broilers experimentally infected with inclusion body hepatitis (IBH) virus. It is presumed that that virus replicating in intestinal tract reached the blood stream via draining lymphatics and thoracic ducts. Chickens do not have discrete lymphnodes as do mammals; there is irregular distribution of a rich, accumulative, diffuse lymphoid tissue (analogous to Peyer’s patches) along the alimentary tract from pharynx to cloaca. This is in addition to the most concentrated gut associated lymphoid tissues which occurs in caecal tonsils and bursa of Fabricius (Toivanen and Toivanen, 1983; Toivanene et al., 1981). Chicken also has a system of lymphatic vessels that collect fluid from tissues and pass it first into a pair of thoracic ducts and then into superior vena cava (Payne, 1971). Small particles, such as adenoviruses, may be taken up from the intestinal lumen by absorptive cells which utilize tubulo–vaculor apparatus (Worthington and Graney, 1972). This process is independent to virus infecting and replicating within the cell. This process may have contributed to the early appearance of viraemia in these birds.
At day 1 PI, the virus was detected only in two tissues (intestine and caecum) from orally infected chickens. Then at day 2PI, it spread to liver, kidney, intestine, bursa of Fabricius, spleen and caecum. Severity of infection was more in liver and kidney of i.m. than orally infected chickens. At day 3 PI, the viral antigens were detected only in the liver, kidney, bursa of Fabricius and spleen. Apart from liver and kidneys, lymphoid organs viz., spleen, thymus and bursa of Fabricius and caecal tonsils are other organs of predilection for hydropericardium syndrome virus and virus from lymphoid organs was capable to cause disease when inoculated in healthy chickens (Naeem et al., 1995). This predilection of virus also explains its immunosuppressive effect.
Only kidney and bursa of Fabricius showed presence of viral antigens at day 5 PI, while at day 10 PI, only kidney, spleen and caecum from chicks infected via i.m. route exhibited presence of viral antigens, while viral antigens could be detected only in spleen of chicks infected via oral route. These findings corroborate the assumption that lymphoid organs are sites of predilection for HPS virus and also suggest that probably kidneys remain infected for longer period than liver. Findings of present study also suggest that for establishing infection in experimental chicks, i.m. route is superior to oral route. Adenoviruses are acid resistant; therefore, they are capable of surviving passage through the stomach and infecting the intestinal epithelium. Kohn (1962) also reported that alimentary tract rather than respiratory tract is the main site of aviadenovirus (AAV) replication. Saiffudin and Wilks (1991) studied the pathogenesis of inclusion body hepatitis by ELISA and immunohistochemistry. Viral antigens were detected from 12 hr to 13 day PI and in the blood plasma by 24 hr PI. Antigens were first detected in the liver from day 2 and reached peak levels by day 6 PI. Initially, viral antigens in the liver were restricted to cells lining the sinusoids but increasing involvement of hepatocytes occurred with time. Small amounts of viral antigen were detected in other tissues. Cook (1983) demonstrated distribution of AAV in different tissues including blood but did not demonstrate when a particular organ was first infected. Kawamura et al. (1963) also demonstrated viremia at 4th DPI and dissemination of virus in the body following AAV–1 (strain Ote) infection.
It is evident from our findings that the most organs were infected between day 3 and 5 PI and mean score of antigen concentration was also high during the period. Cook (1983) also reported that the incubation period of IBH was 3 to 8 days. During this period virus spreads to and rapidly multiplies to target organs including intestine and liver, and produces clinical disease. Antibody titer and severity of infection has inverse relationship as the antibody titer increased severity of infection decreased and lesser number of organs revealed the presence of HPSV antigens. It can also be concluded from our findings that i.m. route is better than oral route for production of hyperimmune serum against HPS virus. Saiffudin and Wilks (1991) reported that VN antibodies in birds experimentally infected with Inclusion body hepatitis (IBH) virus were first detected in blood at 7 days and increased to high titers (mean titer 448) by 13th DPI when viral antigens were undetectable in most organs. It shows a direct relationship between appearance of antibodies and recovery from infection with HPS virus.
COMPETING INTEREST STATEMENT
The authors declare there is no competing interest associated with this study.
ACKNOWLEDGEMENTS
The facilities and financial assistance provided by Dean, College of Veterinary Sciences and Director, Experiment Station, G.B. Pant University of Agriculture & Technology, Pantnagar and Indian Council of Agricultural Research, New Delhi, respectively, are duly acknowledged.
REFERENCES
Abdul-Aziz TA and Hasan SY (1995). Hydropericardium syndrome in broiler chickens: its contagious nature and pathology. Res Vet. Sci. 59: 219–221.
http://dx.doi.org/10.1016/0034-5288(95)90005-5
Abe T, Nakamura K, Tojo H, Mase M, Shibahara T, Yamaguchi S and Yuasa N (1998). Histology, histochemistry and ultrastructure of hydropericardium syndrome in adult broiler breeders and broiler chicks. Avian Dis. 43: 606–612.
http://dx.doi.org/10.2307/1592690
Anjum AD, Sabri MA and Iqbal Z (1989). Hydropericarditis syndrome in broiler chicks in Pakistan. Vet. Rec. 124:247–248.
http://dx.doi.org/10.1136/vr.124.10.247
PMid:2711581
Asrani RK, Gupta BK, Sharma SK, Singh SP and Katoch RC (1997). Hydropericardium hepatopathy syndrome in Asian poultry. Vet. Rec. 141:271–273.
http://dx.doi.org/10.1136/vr.141.11.271
PMid:9316243
Asthana M, Chandra R, and Kumar R (2013). Hydropericardium syndrome: current state and future developments. Archiv. Virol. 158 (5):921–933.
http://dx.doi.org/10.1007/s00705-012-1570-x
PMid:23242777
Calnek BW and Hitchner SB (1969). Localization of viral antigen in chickens infected with Marek's disease Herpes virus. J. Natl. Cancer Inst. 43(4): 935–949.
PMid:4310100
Chandra R, Shukla SK, Kumar M and Garg SK (1997). Electron microscopic demonstration of an adenovirus in the hepatocytes of birds experimentally infected with hydropericardium syndrome. Vet. Rec. 140(3):70–71.
http://dx.doi.org/10.1136/vr.140.3.70-b
PMid:9023909
Choi KS, Kye JY, Kim WJ, Jeon EK, Lee KY and Sung HW (2012). Epidemiological investigation of outbreak of adenovirus infection in commercial chickens in Korea. Poult. Sci. 90(10): 2502–2506.
http://dx.doi.org/10.3382/ps.2012-02296
PMid:22991534
Cook JKA (1983). Fowl adenoviruses: studies on aspects of pathogenicity of six strains for 1–day–old chicks. Avian Pathol. 12(1):35–43.
http://dx.doi.org/10.1080/03079458308436147
PMid:18766761
Fasina SO and Fabricant J (1982). Immunofluorescence studies on early pathogenesis of haemorrhagic enteritis virus infection in turkeys and chicken. Avian Dis. 26(1): 158–163.
http://dx.doi.org/10.2307/1590034
PMid:6284108
Fraser CM, Bergeron JA, Mays A and Aiello SE (1991). Cardiac insufficiency and failure. In: The Merck Veterinary Manual, (Merck & Co., Inc. Rahway, NJ, 46–49. Kawamura H, Sato T, Tsubahara H and Isogai S (1963). Isolation of CELO virus from chicken trachea. Natl. Inst. Anim. Hlth. Q3:1–10.
Kohn A (1962). Gallus adeno–like virus in chicken – studies on infection, excretion and immunity. Am. J. Vet. Res. 23:562–567.
PMid:14457839
Kumar R and Chandra R (2009). Isolation and characterization of fowl adenovirus associated with hydropericardium syndrome. Indian Vet. J. 86(10):1084–1086.
Kumar R, Shukla SK, Chandra R and Agrawal, DK (2003a). An outbreak of classical inclusion body hepatitis in domestic fowl: Isolation and characterization of etiological agent. Indian J. Anim. Sci. 73 (5):477–480.
Kumar R, Chandra R and Shukla SK (2003b). Isolation of etiological agent of hydropericardium syndrome in chicken embryo liver cell culture and its serological characterization. Indian J. Exptl Biol. 41:821–826.
PMid:15248478
Kumar R, Chandra R, Shukla SK. and Rao VDP (2004). Pathological and electron microscopic studies in broiler chicks naturally infected with hydropericardium syndrome virus. Indian J. Vet. Path. 28(2):91–93.
McFerran JB and Adair BM (2003). Group I adenovirus infections. In: Saif YM, ed. Diseases of Poultry. Ames, Iowa: Iowa State Press, 214–227.
Meulemans G, Boschmans M, Van den Berg TP and Decaesstecker M (2001). Polymerase chain reaction combined with restriction enzyme analysis for detection and differentiation of fowl adenoviruses. Avian Pathol. 30(6):655–660
http://dx.doi.org/10.1080/03079450120092143
Naeem K, Niazi T, Malik SA and Cheema AH (1995). Immunosuppressive potential and pathogenecity of an avian adenovirus isolate involved in hydropericardium syndrome in broilers. Avian Dis. 39(4):723–728.
http://dx.doi.org/10.2307/1592408
PMid:8719205
Nakamura K, Mase M, Yamaguchi S, Shibahara T and Yuasa N (1999). Pathologic study of specific–pathogen–free chicks and hens inoculated with adenovirus isolated from hydropericardium syndrome. Avian Dis. 43:414–423.
http://dx.doi.org/10.2307/1592638
PMid:10494409
Niazi AK, Khan MZ and Siddique M (1989). Haematological studies on naturally occurring hydropericardium syndrome in broiler chicks. Vet. Rec. 125(15): 400–403.
http://dx.doi.org/10.1136/vr.125.15.400
PMid:2815524
Payne LN (1971).The lymphoid system. In: Bell, D. J., Freeman, B. M. (eds.) Physiology and Biochemistry of domestic fowl, Vol. 2, Academic Press, London, pp.985–1037.
Ravikumar, Chandra R, Shukla SK, Agrawal DK and Kumar M (1997). Hydropericardium syndrome in India: A preliminary study on causative agent and control of disease by inactivated autogenous vaccine. Trop. Anim. Hlth. Prod. 29(3):158–164.
http://dx.doi.org/10.1007/BF02633014
Saiffudin M and Wilks CR (1990). Development of an Enzyme linked immunosorbent assay to detect and quantify adenovirus in chicken tissues. Avian Dis. 34(2):239–245.
http://dx.doi.org/10.2307/1591404
Saiffudin M and Wilks CR (1991). Pathogenesis of acute viral hepatitis: inclusion body hepatitis in the chicken. Archiv. Virol. 116(1–4): 33–43.
http://dx.doi.org/10.1007/BF01319229
Toivanen A and Toivanen P (1983). The avian immune system In: Disease control and prevention in poultry production. The International Union of Immunological Societies. Proceedings No. 66. The Post Graduate Committee in Veterinary Science, University of Sydney in association with Australian Veterinary Poultry Association, Sydney, pp. 115–122.
Toivanen A, Toivanen P, Eskola J and Lassila O (1981). Ontogeny of the chicken lymphoid system. In: Rose, M. E., Payne, L. N., Freeman, B. M. (eds.) Avian Immunology. British Poultry Science Ltd., Edinburgh, pp. 45–62.
Worthington BB and Graney DO (1972). Uptake of adenovirus by intestinal absorptive cells of the suckling rats. Anat. Rec. 175(1):37–76.
http://dx.doi.org/10.1002/ar.1091750105
PMid:4345520

