
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 1 (3): 96 – 101Molecular Epidemiology and Characterization of Salmonella Serovars from Broilers in Haryana, India
Tarun Kumar1, Nand Kishor Mahajan2, Naresh Kumar Rakha2
- Astt. Technical manager, Bovian health care Pvt. Ltd, 2Professor, Department of Veterinary Public Health & Epidemiology, LLRUVAS, Hisar, Haryana
- Professor, Department of Veterinary Medicine, LLRUVAS, Hisar, Haryana, India
*Corresponding author:tarunvet@gmail.com
ARTICLE CITATION:
Kumar T, Mahajan NK and Rakha NK (2013). Molecular epidemiology and characterization of Salmonella serovars from broilers in haryana, india. Adv. Anim. Vet. Sci. 1 (3): 96 – 101.
Received: 2013–07–29, Revised: 2013–08–03, Accepted: 2013–08–04
The electronic version of this article is the complete one and can be found online at
(
http://www.nexusacademicpublishers.com/table_contents_detail/4/66/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
A total of 51 isolates of Salmonella, including 23 isolates of Salmonella gallinarum, 13 isolates of Salmonella pullorum, 9 isolates of Salmonella enteritidis and 6 isolates of Salmonella typhimurium were isolated and characterized from outbreaks of Salmonella infection in broilers from commercial poultry farms of Haryana state, India. Isolates were characterized at their genotypic level by plasmid profiling, restriction endonuclease analysis and PCR. Plasmid profiles and antibiotic resistance pattern were correlated. Eight different plasmid profiles were obtained with presence of large plasmid of size 85 kb in maximum number of isolates (70%). Study suggested that 85 kb plasmid might carry the genes responsible for virulence. No positive correlation between plasmid profiles and antibiotic resistance pattern was found. Restriction endonuclease analysis of genomic DNA showed a high degree of genome homogeneity suggesting a common grandparent source of infection. PCR was found to be sensitive, specific and fast method to know the virulence of Salmonella isolates. Study concluded that plasmid profiling along with restriction enzyme analysis can be used as epidemiological markers in back tracing infections especially in case of outbreaks. Need of hour is to start mandatory Salmonella testing of parent flocks, positive reactors should be removed from the flock and vaccination of parent flocks should be carried out for control of this fast spreading, zoonotically important vertically transmitted pathogen. Moreover, at government level, there should be some legislation for control of such infections from hatchery.
INTRODUCTION
Bacterial infections of genus Salmonella are responsible for large number of diseases in poultry. These diseases cause significant economical loss to poultry farmers. Moreover infected poultry stocks are the main reservoirs of Salmonella that can be transmitted to humans through food chain (Joseph et al., 1997). Since the year 2005, there is sudden increase of Salmonella infection in poultry flocks mainly broilers in Haryana, India (Kumar et al., 2010). To explore reasons for this increasing trend of Salmonella infection, molecular epidemiology at genotypic level was conducted. Molecular epidemiology is the application of techniques of molecular genetics to type strains of infectious agents for epidemiological studies. Detailed strain identification is essential for successful epidemiological investigation of Salmonella enterica serovar outbreaks. Investigations have relied traditionally on serological methods antibiograms, Phage typing etc. Modern typing methods are based on characterization of the genotype of the organism. In this regard Plasmid profiles have become valuable markers in the epidemiological investigation (Ferris et al., 1992; Joseph et al., 1997).
Plasmid profile analysis has been used as a rapid method and has shown success in the discrimination of Salmonella strains (Taylor et al., 1982). Characterization of Salmonella enterica serovar gallinarum and pullorum was performed by plasmid proofing and biochemical analysis (Christensen et al., 1992). Search for plasmid associated virulence gene is of great epidemiological interest and Josh and his co–workers in 1999 detect this gene by PCR. Polymerase chain reaction (PCR) can particularly be a useful tool to provide rapid and definitive detection of avian Salmonella serotypes. Strains having 85 Kb plasmid proved to be virulent (Joseph et al., 1999). An association between the presence of plasmid of an approximately 85–kilobase (kb) in Salmonella enterica serovar gallinarum and ability of strains to produce high mortality in chickens was reported (Barrow et al., 1987). Epidemiological status of avian Salmonella enterica serovar infections in India was elucidated (Prakash et al., 2005). The restriction endonuclease analysis of total DNA was found to be useful in differentiating field isolates of Salmonella enterica serovar gallinarum (Joseph and Singh 2000). Antibiogram study and plasmid profile analysis were conducted to find out the correlation of recently isolated Salmonella strains of Bangladesh (Khan et al., 2005). In India, although the isolation of various Salmonella serovars from poultry has been regularly reported from different parts of the country but information about the molecular characteristics of these serovars is limited (Mariani et al., 2001). Moreover there are very few reports to date on genotypic analysis of Salmonella spp. isolated from poultry in this region too.
In the present study, isolation was made from suspected outbreaks, attempts were made to find out the pattern of plasmids in isolates from different sources and find out the differences /relatedness to derive an epidemiological pattern at genotypic level of this infection in the region so as to recommend preventive methods for this economically important zoonotic disease of poultry.
MATERIALS AND METHODS
Sampling and isolating Salmonella
In the year of 2008 a total of 132 outbreaks were recorded from various poultry farms (mainly broilers) situated in Haryana, India. Samples were collected for isolation and 51 isolates of Salmonella, including 23 isolates of Salmonella enterica serovar gallinarum, 13 isolates of Salmonella pullorum, 9 isolates of Salmonella enteritidis and 6 isolates of Salmonella typhimurium were isolated from these outbreaks. Initially isolates were confirmed by culture characteristics and biochemical assay with the help of Hi Salmonella Identification kit (Himedia). Finally serotyping was performed at National Salmonella and Escherichia Centre, Central Research Institute, Kasauli – 173204, Himachal Pradesh, India. Isolates were also characterized by scanning electron microscopy. Molecular characterization was performed by plasmid profiling, restriction enzyme analysis, PCR. The in vitro antimicrobial sensitivity patterns of Salmonella isolates to various antimicrobial agents, Chloremphenicol C (30 mcg), Amikacin Ak (30 mcg), Gentamicin G (10 mcg), Cephatoxime Ce (10 mcg) and Cephadroxil Cq (30 mcg) were determined by the disc diffusion technique (Bauer et al., 1996).
PLASMID PROFILING
Plasmid DNA isolation
Plasmid DNA was extracted using alkaline lysis method (mini prep) as described in Sambrook and Russel 2001. Briefly single colony of Salmonella spp. was inoculated in 3 ml BHI broth and was incubated at 37¬oC for 16–18 hours with intermittent shaking. Out of this, 1.5 ml was centrifuged at 8200 X g for 5 minutes at 4oC. Supernatant was removed. Bacterial pellet was resuspended in 100ul of ice cold alkaline lysis solution I by vigorous vortexing and incubated at 25oC. Two hundred ul of freshly prepared alkaline lysis solution II was added to bacterial suspension and was mixed. Tube was kept on ice for 3–5 minutes. One hundred fifty ul of ice cold alkaline lysis solution III was added and dispersed through the viscous bacterial lysate by inverting the tube several times and tube was kept on ice for 3–5 minutes. Bacterial lysate was centrifuged at 8200 X g for 10 minutes at 4oC. Equal volume of phenol: chloroform: isoamyalcohol (25:24:1) was added in the Supernatant. Organic and aqueous phases were mixed by vortexing and then the emulsion was centrifuged at 8200 X g for 10 minutes at 4oC. Aqueous upper layer was transferred to a fresh tube and equal volume of chloroform: isoamyalcohol (24:1) was added to remove the traces of phenol. Organic and aqueous phases were mixed by vortexing and then the emulsion was centrifuged at 8200 X g for 10 minutes at 4oC. Aqueous upper layer was transferred to a fresh tube. Nucleic acids were precipitated from the supernatant by adding equal volume of isopropanol and incubated at –20oC for 15 minutes. Precipitated nucleic acids were collected by centrifugation at 8200 X g for 20 minutes at 4oC. Supernatant was removed by gentle aspiration. Pellet was resuspended in 1 ml of 70% ethanol. DNA was recovered by centrifugation at 8200 X g for 20 minutes at 4oC and supernatant was removed by gentle aspiration. Pellet was dried in the desiccators concentrated. Nucleic acids were dissolved in 20ul of tris EDTA (TE) (PH 8.0) containing 20 ug / ml DNase– free RNase A solution. DNA was stored at – 20oC till further use.
Agarose gel electrophoresis
The isolated plasmids were separated in 0.7% agarose gel (Low LEO) in 0.5X Tris Borate EDTA (TBE) electrophoresis buffer. Ethidium bromide, 0.5 ug / ml (Sigma) was added in the gel and photograph was taken using digital camera (Canon Powershot A620) under UV transilluminator (Spectroline Biovision, USA) after electrophoresis. Molecular weight/size marker ranging from 125bp to 23.13kbp was run along with isolated plasmid DNA. Standard curve between log molecular size of DNA marker and its Rf. values was plotted and point to point method was used to calculate molecular size of plasmids.
Restriction enzyme analysis
Isolation of chromosomal DNA
Bacterial chromosomal DNA was isolated by the method outlined by Maloy et al., 1996 with certain modifications. Briefly, Salmonella isolate was grown in 300 ml of BHI broth, pelleted by centrifugation, washed twice in PBS and then in T50E50(50 mM Tris–HCL and 50 mM EDTA, pH 8.0), the bacteria were finally pelleted and resuspended in 10 ml of T50E50 in 50 ml sterile centrifuge tube. In the tube was added 1.250 ml of T 50E50 containing 20 mg/ml of freshly prepared lysozyme. Tubes were incubated for 30 minutes at RT. Then 250ul of SDS was added in the tube followed by 1.250 ml T 10E1 (10 mM Tris–HCI and 1 mM EDTA, pH 7.4) containing Pronase E (500 µg/ml stock in DGDW). The mixture was incubated at 37°C for 1.5 hours with intermittent shaking. Equal volumes of tris–HCI buffered phenol was then added and mixed by gentle rocking for 3 minutes and incubated at 37°C for 1 hour. The tubes were centrifuged at 3000 x g for 10 minutes at RT. Upper aqueous phase was collected into a clean centrifuge tube. The aqueous phase collected, was extracted twice with phenol: chloroform: isoamyl alcohol (25: 24: 1) by mixing 1: 1. The centrifuge tube was spun for 1 minute at RT and the upper phase was collected. To this 3M sodium acetate was added to a final concentration of 0.15 M. The DNA was precipitated by adding 2 volumes of ice–cold absolute alcohol. The precipitated DNA was collected by spooling on a sealed micropipette tip. Twirling of the sealed pipette tip released the spooled DNA in a tube containing 4 ml of T10E1. It was stored overnight at 4°C in a refrigerator. To the DNA preparation was added 200 µl of 10 mg/ml stock solution of RNase. After this, 3M sodium acetate to a final concentration of 0.15 M, was added and the DNA was precipitated with 2 volumes of cold ethanol. The precipitated DNA was spooled on a sealed pipette tip, pulled out on the tip, rinsed with 70% ethanol in another tube, air dried in the same tube and stored at –200 C. Further purification of the semipurified DNA was achieved by repeated phenol–chloroform treatment. Finally, the purified DNA was resuspended in endotoxin–free water and stored at –20oC.
Restriction enzyme analysis
Purified DNA from three field isolates (Salmonella gallinarum) of different outbreaks were subjected to RE analysis with two enzymes namely HindIII, EcoRI (Fermentas) to find out the genomic linkage. Restriction enzymes namely HindIII, EcoRI, PStI , Ssp1and Bstx1 (Fermentas) were also used to know which enzyme give interpretable banding patterns for typing Salmonella gallinarum (T16). After digestion, all the samples were analyzed on 2.0 % agarose gel with 23 kb DNA ladder.
Polymerase chain reaction (PCR)
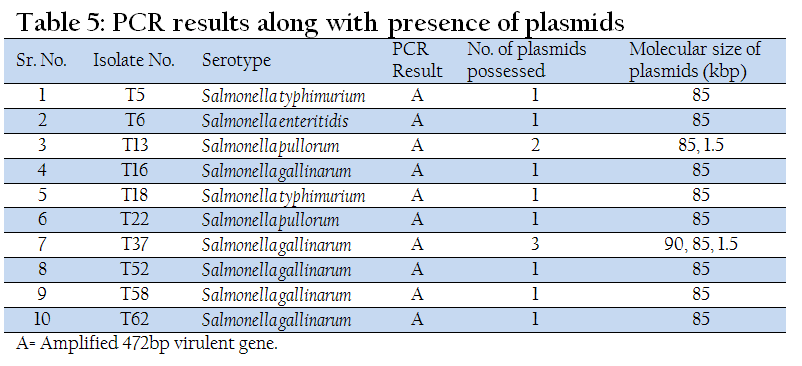
Ten field isolates of Salmonella including 5 isolates of Salmonella gallinarum, 2 isolates of each Salmonella pullorum and Salmonella typhimurium and one isolate of Salmonella enteritidis were subjected for PCR to amplify plasmid (85 kb) associated virulence gene as described earlier (Rexach et al., 1994). The details of primers (sigma) used are as follows:
Gene |
Primers |
Primer sequence (5’–3’) |
Virulence–associated plasmid gene |
Forward |
TTG TAG CTG CTT ATG ATG GGG GGG |
Reverse |
TGG AGA AAC GAC GCA CTG TAC TGC |
Two microliters of the prepared DNA template was added to 25 ml of PCR reaction mixture in 0.2 ml thin walled microfuge tube. The reaction mixtures used in the PCR steps contained 10 X PCR buffer, 25 mM MgCl 2 , 10 mM deoxynucleoside triphosphate, 1.0 µl of forward primer, 1.0 µl of reverse primer (as indicated above), and 0.5 µl of Taq DNA polymerase. Plasmid DNA amplification was carried out in a Eppendorf thermocycler using an initial denaturation step of 94 oC for 4 min, followed by 32 cycles of amplification with denaturation at 94 oC for 50 s, annealing at 64 oC at for 60 s, and extension at 72 oC for 60 s, ending with a final extension at 72 oC for 5 min. Upon completion of PCR, the products were analyzed by ethidium bromide stained 1% agarose gel electrophoresis
RESULTS
Plasmid profiling
Among 23 isolates of Salmonella gallinarum, 21 isolates (91.30%) possessed plasmids of different molecular weights and only two isolates (8.70%) were without plasmid. A common 85 kb large plasmid was present in 20 isolates and only one isolate was devoid of this plasmid. Small plasmids of size 1.5 kb and 2.5 kb were also present. Six isolates had only one plasmid of size 85 kb, while one isolate had one plasmid of size 1.5 kb alone. Rest of the isolates (fourteen) showed presence of multiple plasmids. Plasmid profile analysis was able to subtype the 23 isolates of Salmonella gallinarum into seven groups.
Eleven out of thirteen isolates of Salmonella pullorum contained plasmids of different molecular weights. Five isolates showed the presence of single large plasmid of size 85 kb, while three isolates possessed small plasmid of size 1.5 kb. A 2.5 kb plasmid was detected in one isolate (T35). Two isolates showed the presence of both large and small plasmids of size 85 kb and 1.5 kb. Six different plasmid profiles were obtained in serovar gallinarum while four plasmid profiles were seen in serovar pullorum.
In Nine isolates of Salmonella enteritidis two isolates showed presence of 2 large plasmids of size 90 kb and 85 kb. Three isolates possessed only one large plasmid of size 85 kb. In one isolate both 85 kb and 1.5 kb plasmids were present. Overall four different plasmid profiles were present.
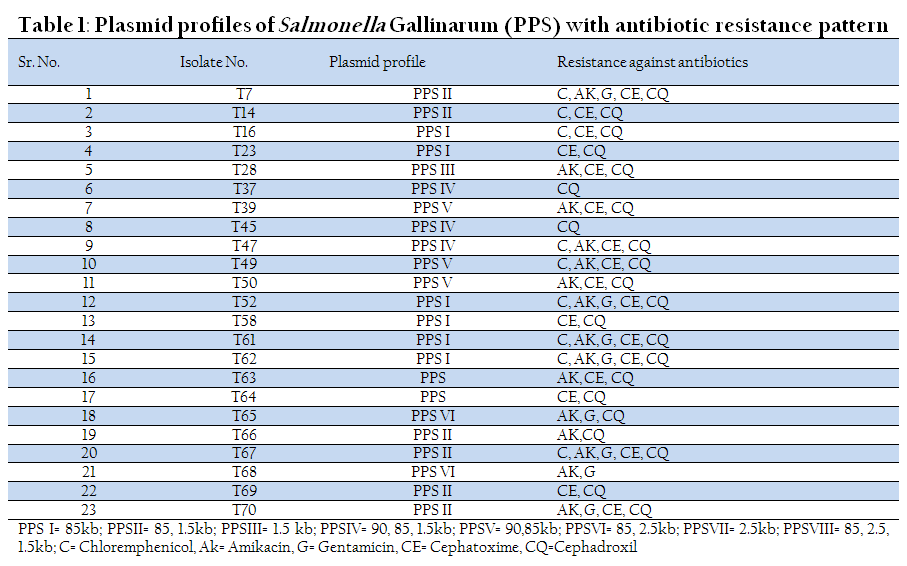
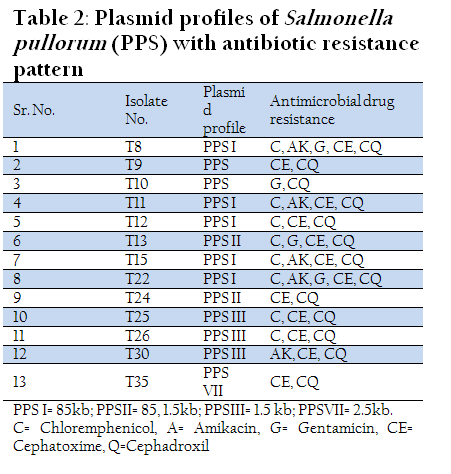
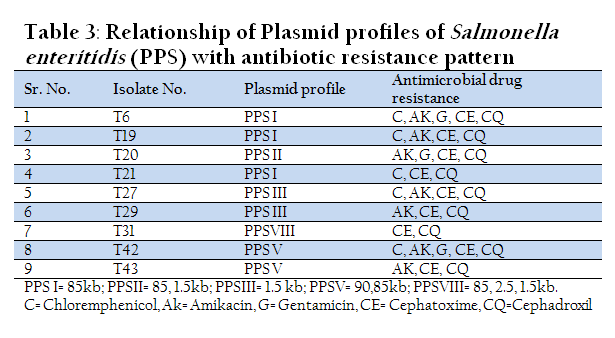
Table 3: Relationship of Plasmid profiles of Salmonella enteritidis (PPS) with antibiotic resistance pattern
In another zoonotically important serotype Salmonella typhimurium out of six isolates, 4 isolates possessed only large plasmid of size 85 kb. Two isolates were devoid of any plasmid (Table 1, 2, 3, &4 and Figure 1(a and 1(b).
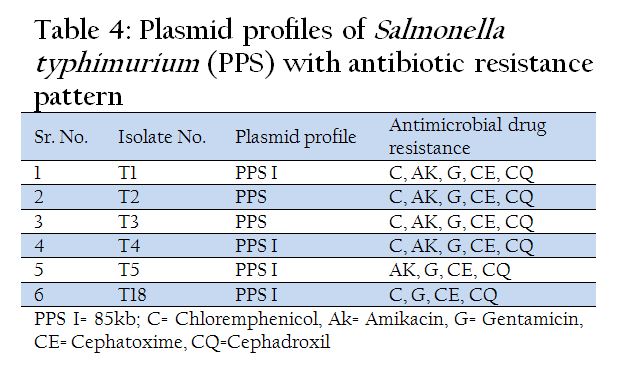
All the isolates of Salmonella spp. showed multiple drug resistance patterns. Salmonella gallinarum and Salmonella enteritidis isolates showed maximum sensitivity to Chloramphenicol. Gentamicin was found to be the second most effective antibiotic. Salmonella pullorum showed maximum sensitivity to Gentamicin (62.5%), where as Salmonella typhimurium isolates were 100% resistant to all antibiotics used in the study except Chloremphenicol which showed sensitivity for 33.4% isolates. Both Salmonella enteritidis and Salmonella typhimurium are zoonotically important serotypes and these isolates showed maximum resistance to commonly used antibiotics which is a cause of concern. However, there was no positive correlation between presence of plasmid and drug resistance pattern.
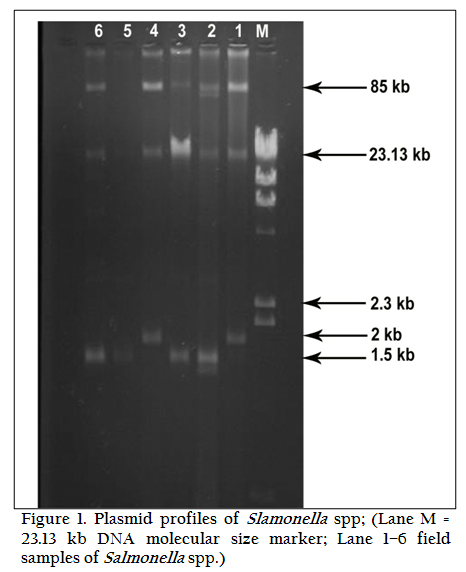
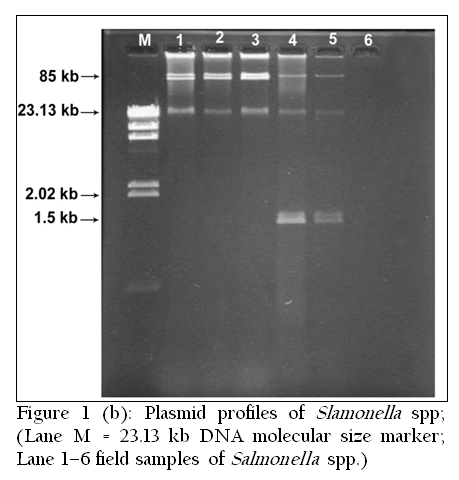
Restriction enzyme endonuclease analysis
Bacterial genomic DNA of 3 Salmonella gallinarum isolates having purity 1.8 by Biophotometer showed complete digestion of genomic DNA with large number of bands. Similar banding pattern was seen in all the three isolates establishing the fact that these were similar at genomic level. When these isolates were analyzed for plasmids, it was observed that they had only single plasmid of the size 85 kb. These findings suggested that the isolates were similar at genomic level and might have originated from the single grandparent source. Findings were supported by retrospective study which revealed that most of the affected farms receive the poultry birds from a single hatchery. Digestion of Salmonella gallinarum (T16) with EcoRI, HindIII and PstI showed similar banding pattern with almost all the bands of high molecular weight. With EcoRI and HindIII last band was of the size approximately 1 kb. When banding pattern with BstXI and SspI was analysed, more discrete bands of lower molecular weight were observed. With BstXI last band was of the size approximately 100bp while with SspI, it was nearly of 150 bp.
Polymerase chain reaction (PCR)
All the ten isolates were having one common plasmid of size 85 kb and specifically amplified a product of approximately 472 base pair (bp), showing the presence of plasmid gene linked to virulence in Salmonella irrespective of the serotype (Table 5, Figure 2). PCR detection of plasmid–associated virulence gene, presence of 85 kb plasmid and high mortality in one to two week old chicks were positively correlated.
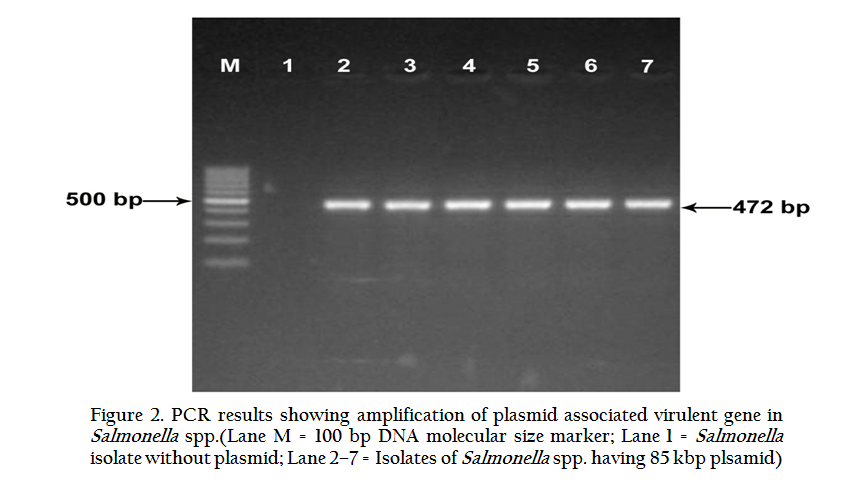
DISCUSSION
Detailed strain identification is essential for successful epidemiological investigation of Salmonella enterica serovar outbreaks. Plasmid profile analysis has been used as a rapid method and has shown success in sub–typing of Salmonella strains. Plasmids often encode properties such as antimicrobial resistance, invasiveness capability, toxins etc. Analyses on the bases of plasmid profiling revealed that in a given geographic area, an individual bacterial clone predominates (Van Embden et al., 1976; Helmuth et al., 1981). In Britain, plasmid profiling was first applied to know Salmonella epidemiology in 1979. Large plasmids of various sizes have been found in several serotypes of Salmonella, all of which produce a systemic type of disease in animals or humans. The role of 60–megadalton plasmid in the virulence of Salmonella typhimurium was demonstrated (Jones et al., 1982). An association between presence of 85–kilobase plasmid in Salmonella gallinarum and ability of strains to produce high mortality in chickens was reported earlier (Barrow et al., 1987).
In the present study Salmonella isolates were characterized at their genotypic level by plasmid profiling, restriction enzyme analyses and PCR. Six different plasmid profiles were obtained in serovar gallinarum while four plasmid profiles were seen in serovar pullorum. A common large plasmid of size 85 kb was present in majority of the isolates along with the presence of small plasmids of size 1.5 kb and 2.5 kb. Presence of more than one plasmids detected in the present study is in agreement with the earlier observations (Rahman, 1999). Correlation between plasmid profiles and antibiotic resistance found to be useful in epidemiological characterization of Salmonella isolates (Bhattacharya et al., 2001; Radu et al., 1997). But in the present study there was no positive correlation between plasmid profiles and antibiotic resistance pattern. The results of the present study point towards the possibility of drug resistance–genes being present on the chromosomal DNA, in addition to the plasmids, since isolates which were devoid of plasmids also showed resistance to multiple antibiotics. However presence of plasmids correlated with virulence factors was previously reported (Oh et al., 2002; Singh et al., 1996) but various workers reported that drug resistance genes may also be present on chromosomes (Mariani et al., 2001). Multiple drug resistance was present in most of the isolates which might also be because of the presence of large number of plasmids in the isolates carrying drug resistance genes (Bakshi et al., 2003).
Plasmid profiling analyses along with restriction endonuclease analysis revealed that these isolates showed a high degree of genome homogeneity and might have originated from the single grandparent source. Digestion of genomic DNA of Salmonella gallinarum with different restriction enzymes showed similar banding pattern with almost all the bands of high molecular weight. These observations conform to the observations of Tompkins et al. (1986). However BstXI and SspI yielded more discrete bands of lower molecular weight.
Plasmid associated virulence gene was detected by PCR irrespective of the serotype. Various other workers (Bhattacharya et al., 2001; Mariani et al., 2001) also detect Salmonella virulence–associated plasmid gene by amplifying 472 bp product. In the present study one common plasmid of 85 kb size was present in all the isolates which were positive by PCR. Retrospective study showed that the isolates with 85 kb plasmid were responsible for high mortality in the affected flock, suggesting the association of virulent gene with the presence of 85 kb plasmid. This is corroborated by the findings of other workers (Bhattacharya et al., 2001; Joseph et al., 2000; Rexach et al., 1994).
CONCLUSIONS
Study concluded that plasmid profiling along with restriction enzyme analysis can be used as epidemiological markers in back tracing infections especially in case of outbreaks. As 75% of isolates possess 85 kb plasmid, there is some common source of infection possibly at grandparent level. No positive correlation between plasmid profiles and antibiotic resistance pattern was found. The multi–drug resistance of the isolates suggests indiscriminate use of antimicrobials in the developing countries like India. PCR proved to be sensitive and effective method to detect virulent gene of Salmonella infection. So need of hour is to start mandatory Salmonella testing of parent flocks, positive reactors should be removed from the flock and vaccination of parent flocks may be carried out for control of this fast spreading, zoonotically important vertically transmitted pathogen. Moreover at government level there should be some legislation for control of such infections from hatchery.
ACKNOWLEDGEMENT
Director and laboratory staff of the National Salmonella and E. Coli, Central Research Institute, Kasauli– H.P, for serogrouping the Salmonella isolates.
REFERENCES
Bakshi CS, Singh VP, Malik RK, Singh RK and Harma B (2003). 55 kb plasmid and virulence associated genes are positively correlated with Salmonella enteritidis pathogenicity in mice and chickens. Vet. Res. Communication. 27: 425–432.
http://dx.doi.org/10.1023/A:1025720306045
PMid:14582741
Barrow PA, Simpson JM, Lovell MA and Binns MM (1987). Contribution of Salmonella gallinarum large plasmid toward virulence of fowl typhoid. Infection and Immunity. 55: 388–92.
PMid:3804442 PMCid:PMC260339
Bauer AW, Kirby WMM, Sherries JC and Truck M (1996). Antibiotic susceptibility testing by standardized single disc method. American Journal of Clinical Pathology. 45: 493.
Bhattacharya A, Singh VP and Verma JC (2001). Phenotypic and genotypic characterization of Salmonella typhimurium isolates of avian origin. Indian Journal of Animal Health. 40: 123–128.
Christensen JP, Olsen JE, Hansen HC and Bisgaard M (1992). Characterization of Salmonella enterica serovar gallinarum biovars gallinarum and pullorum by plasmid profiling and biochemical analysis. Avian Pathology. 22: 725–738.
http://dx.doi.org/10.1080/03079459308418960
PMid:18671057
Ferris KE, Andrews RE, Theon CO and Blackburn BO (1992). Plasmid profile analysis, phage typing and antibiotic sensitivity of Salmonella dublin from clinical isolates in the United States. Veterinary Microbiology. 32: 51–62.
http://dx.doi.org/10.1016/0378-1135(92)90006-F
Helmuth R, Stephan R, Bulling E,Van Leeuwen WJ, Van Embden JD, Guinee D, Portnoy D and Falkow S (1981). R–factor cointegrate formation of Salmonella typhimurium bacteriophage type 201 strains. J. Bacteriol. 146: 444–452.
PMid:7012128 PMCid:PMC216985
Jones GW, Rabert DK, Svinarich DM and Whitfield HJ (1982). Association of adhesive, invasive, and virulent phenotypes of Salmonella typhimurium with autonomous 60–megadalton plasmids. Infect. Immun. 38: 476–486.
PMid:6128304 PMCid:PMC347764
Joseph T, Chaudhuri P, Singh VP and Sharma B (1997). Randomly cloned chromosomal fragment for fingerprinting Salmonella gallinarum isolates. Indian Veterinary Journal. 74: 191–194.
Joseph T, Singh C, Singh VP and Sharma B (1999). Correlation between presence of 85 kb plasmid, amplification of virulence gene and chick pathogenecity in Salmonella gallinarum. Indian Veterinary Journal. 76: 90–93.
Joseph TC and Singh VP (2000). Restriction enzyme analysis of genomic DNA in Salmonella gallinarum field isolates. Indian J Comp Microbiol Immunol Infect Dis. 21 (1): 66–67.
Khan MFR, Rahman MB, Khan MSR, Nazir NH and Rahman M (2005). Antibiogram and Plasmid profile analysis of isolated poultry Salmonella of Bangladesh. Pakistan Journal of Biological Science. 8 (11): 1614–1619.
http://dx.doi.org/10.3923/pjbs.2005.1614.1619
Kumar T, Mahajan NK and Rakha NK (2010). Epidemiology of fowl typhoid in Haryana, India. World's Poultry Science Journal. 66: 503–509,
http://dx.doi.org/10.1017/S0043933910000565
Rexach L, Dilasser F and Fach P (1994). Polymerase chain reaction for Salmonella virulence–associated plasmid genes detection: a new tool in Salmonella epidemiology. Epidemiology and Infection. 112, 33–43.
http://dx.doi.org/10.1017/S0950268800057393
PMid:8119363 PMCid:PMC2271485
Maloy SR, Stewart VJ and Taylor RK (1996). DNA extraction In: The Genetic Analysis of pathogenic bacteria: A laboratory manual. Cold Spring Harbor, Laboratory Press, New York, 408–410.
Mariani P, Barrow PA, Cheng HH, Groenen MM, Negrini R and Bumstead N (2001). Localization to chicken chromosome 5 of a novel locus determining salmonellosis resistance. Immunogenetics. 53 (9):786–91.
http://dx.doi.org/10.1007/s00251-001-0387-7
PMid:11862411
Mir IA, Wani SA, Hussain I, Qureshi SD, Bhat MA and Nishikawa Y (2010). Molecular epidemiology and in vitro antimicrobial susceptibility of Salmonella isolated from poultry in Kashmir. Rev sci tech Off int Epiz. 29 (3): 677–686.
Oh G, Kim S, Lee K, Ha J, Park S, Jung K, Lee K, Song J, Oh GH, Kim SW, Lee KH, Ha JS, Park SC, Jung KS, Lee KW and Song JC (2002). Virulence and plasmid profiles of Salmonella gallinarum isolated from chickens. J Vet Clinics. 19: 159–164.
Prakash B, Krihnappa G, Muniyappa L and Kumar S (2005). Epidemiological characterization of Avian Salmonella enterica serovar infections in India. Int J Poult Sci. 4: 388–395.
http://dx.doi.org/10.3923/ijps.2005.388.395
Radu S, Maimunah M, Rusul G, Mokhtar AM and Khoo LL (1997). Characterization of Salmonella enteritidis isolates by plasmid profiling, phage typing and antibiotic resistance. Trop Biomed. 14: 75–80.
Rahman H. Plasmid profile of Salmonella of animal origin (1999). Indian J Comp Microbiol Immunol Infect Dis. 20: 23–25.
Sambrook J and Russel DW (2001). Molecular Cloning a Laboratory Manual. Cold Spring Harbor, Ny: Cold Spring Labor Laboratory Press, 1: 32–34.
Singh V, Singh VP, Gupta PK and Chaudhuri P (1996). Plasmid profile and virulence analysis of Salmonella gallinarum Indian isolates. J Appl Anim Res. 9: 129–133.
http://dx.doi.org/10.1080/09712119.1996.9706114
Taylor DN, Wachsmuth IK and Shangkuan YH (1982). Salmonella associated with marijuana: a multistage outbreak traced by plasmid fingerprinting. N Engl J Med. 306: 1249–53.
http://dx.doi.org/10.1056/NEJM198205273062101
PMid:7070444
Tompkins LS, Troup N, Labigne–Rousel A and Cohen M (1986). Cloned, random chromosomal sequences as probes to identify Salmonella species. J Infect Dis. 154: 156–62,
http://dx.doi.org/10.1093/infdis/154.1.156
Van Embden JDA, Van Leeuwen WJ and Guinee PAM (1976). Interference with propagation of typing bacteriophage by extrachromosomal elements in Salmonella typhimurium bacteriophage type 505. J Bacteriol. 127: 1414–1426.
PMid:783145 PMCid:PMC232938

