
Advances in Animal and Veterinary Sciences


Short Communication
Advances in Animal and Veterinary Sciences 1 (4): 123 – 126Polymorphism in Catsper1 Gene in Crossbred (Bos Taurus X Bos Indicus) Cattle
Thangavel Geetha, Subodh Kumar*, Prem Prakash Dubey, Balasubramaniam Sivamani, Arjava Sharma
-
Division of Animal Genetics, Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh, 243 122, India
*Corresponding author:subkum@gmail.com
ARTICLE CITATION:
Geetha T, Kumar S, Dubey PP, Sivamani B, Sharma A (2013). Polymorphism in Catsper1 gene in crossbred (Bos taurus X Bos indicus) cattle. Adv. Anim. Vet. Sci. . 1 (4): 123 – 126.
Received: 2013–06–30, Revised: 2013–07–28, Accepted: 2013–07–29
The electronic version of this article is the complete one and can be found online at
(
http://www.nexusacademicpublishers.com/table_contents_detail/4/83/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Nucleotide variability in exon 2 of CatSper1 gene (434 bp) was evaluated in crossbred cattle named Vrindavani (n = 100) using PCR–RFLP and nucleotide sequencing. Using DNA from blood, restriction enzyme analysis with EcoRI and HindIII produced two bands of 315 bp, 119 bp and 360 bp, 74 bp respectively. This amplicon of CatSper1 gene exhibited absence of polymorphism with respect to restriction enzymes used and accordingly, the allelic frequency was found to be unity. The monomorphic pattern of the amplicon of CatSper1 gene with respect to different enzymes indicated the conservedness of this gene. The amplicon was subjected to DNA sequencing, annotation and submitted to GenBank. In order to study the variation at nucleotide level, the sequence of this amplicon was compared with similar sequences of mithun, goat and buffalo and nucleotide variations ascertained.
Semen cryopreservation has been an essential tool to meet the continuously increasing demand of male germplasm in order to sustain the increasing population of crossbreds. The ultimate goal of semen preservation has been to obtain pregnancies after artificial insemination, which would be as effective as natural mating. The successful preservation of semen depends on several factor, which are involved in the process of cryopreservation and the term ‘freezability’ has been given to mean the ability of semen to survive after being frozen without suffering substantial damage. The damage to the semen expressed as sperm count, motility and fertility but practically, it largely refers to post thaw motility (PTM) (Ravimurugan et al., 2007) which again depends on initial/pre freeze motility and is the ultimate trait which qualifies the semen for artificial breeding. Crossbred bulls have high percentage of abnormal spermatozoa, lower level of sperm motility and viability causing decline in fertility rate (Dhanju et al., 2006). Poor sperm motility and freezability of semen has also been reported in Vrindavani crossbred bulls (Ghosh et al., 2007). Researchers have reported more than 50 % rejection rate in different crossbred bulls (Chacon et. al., 1999; Tyagi et. al., 2006; Geetha et al., 2011) due to poor seminal attributes specially sperm motility. Many genes are known to control sperm motility. Off late, an ion channel gene was identified which had a significant bearing on sperm motility (Ren et al., 2001; Quill et al., 2001). This was named as Cation channel of sperm (CatSper) and reported to have four subunits. CatSper family mutations resulted in male infertility (Darszon et al., 2006). CatSpers1–4 are expressed in testis and localized primarily to the principal piece of sperm tail (Ren et al., 2001; Quill et al., 2001; Lobley et al., 2003; Jin et al., 2005). CatSper1 is required for the hyper–activation of sperm cell motility which is essential for fertility (Qi et al., 2007). This study was, therefore, undertaken to ascertain polymorphism of CatSper1 gene in Vrindavani crossbred cattle. This would help in initiating a step in searching the promising DNA markers that could be developed to improve sperm motility of crossbred cattle by assisting in bull selection process.
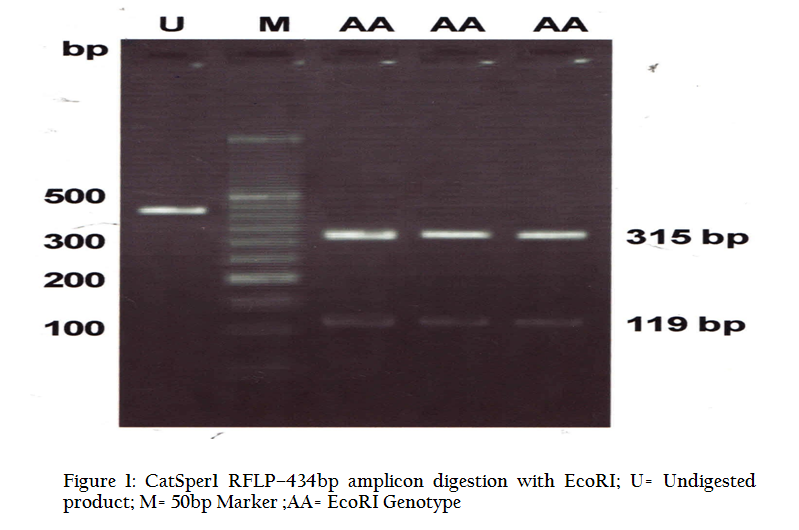
Figure 1: CatSper1 RFLP–434bp amplicon digestion with EcoRI; U= Undigested product; M= 50bp Marker ;AA= EcoRI Genotype
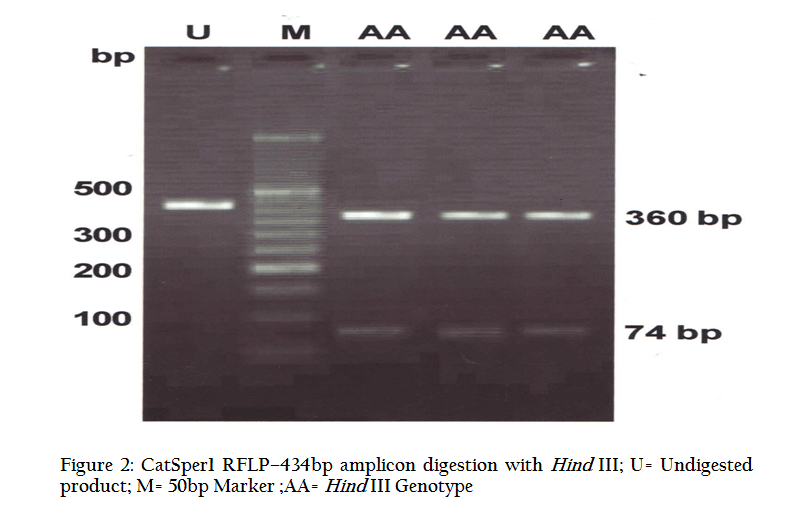
Figure 2: CatSper1 RFLP–434bp amplicon digestion with Hind III; U= Undigested product; M= 50bp Marker ;AA= Hind III Genotype
A total of hundred randomly selected crossbred cattle named Vrindavani (50–62.5 % exotic inheritance comprising of Holstien Frisian, Brown Swiss, Jersey; with Hariana as indigenous stock) developed at the Indian Veterinary Research Institute, Izatnagar and maintained at Livestock Production Management Section (LPM) were included in the present investigation. Venous blood (10 mL) was collected from each crossbred animal in sterile 15 mL polypropylene centrifuge tube containing 0.5 mL of 2.7% EDTA as anticoagulant. DNA isolation was done by phenol chloroform extraction method (Sambrook and Russel, 2001) and the precipitated DNA was dissolved in 200 µL of TE buffer. The quality, purity and concentration of genomic DNA was evaluated by 0.7% agarose gel electrophresis and spectrophotometer (PG Instruments, UK). The samples that showed a clear band (no smearing) in electrophoresis and an OD ratio (OD260/OD280) in the range of 1.7–1.9 were assessed to be of good quality and purity. Their concentration ranged from 650–2100 ng/μL
_112.PNG)
_112.PNG)
A pair of primer was designed by the help of DNASTAR software to amplify CatSper1 gene on the basis of already reported sequence (in silico generated/ predicted) in Bos taurus cattle (NC_007330) available in the GenBank (www.ncbi.nlm.nih.gov). This primer set viz. forward: 5’ GAA GCC CCG TCT GAT GGT TA 3' and reverse: 5' CTG GGA AAG GGA TGT GGA G 3') amplified a 434 bp fragment of CatSper1 gene in the genomic DNA of crossbred cattle consisting of complete exon 2 and flanking introns. The reaction mixture and PCR programme were optimized to achieve the satisfactory level of amplification in a final volume of 25μL containing 1μL genomic DNA (80–100ng), 2.5μL of 10xPCR (1.5mM), 2.5 μL of dNTPs mix (0.2mM), 1.5 μL of MgCl2 (1.5mM), 1μL each of forward and reverse primers (30 pmoles/uL) and 0.2μL of Taq DNA polymerase (5 U/μL). Samples were amplified for 35 cycles (iCycler, Biorad, USA) with initial denaturation at 940C for 3 min., cyclic denaturation at 940C for 1 min., annealing at 55°C for 1min., extension 720C for 1 min. and final extension 720C for 10min. The 434bp amplicon was digested with EcoRI as well as HindIII restriction enzymes which had recognition frames as G↓AATTC and A↓AGCTT, respectively. The digested products were electrophoresed in 2.5 % w/v agarose gel, (stained with ethidium bromide), at 100 V for 5min and then 90V for 1 hr in 1x TBE buffer and visualized under UV light. The amplicon of CatSper1 gene was eluted, cloned in pGEMT vector and sequenced in both orientations. The obtained sequence on crossbred cattle was then aligned with similar sequences in other species viz. zebu cattle (Bos indicus), mithun (Bos frontalis), buffalo (Bubalus bubalis) and goat (Capra hircus) using MEGALIGN module of DNASTAR software (Lasergene, USA).

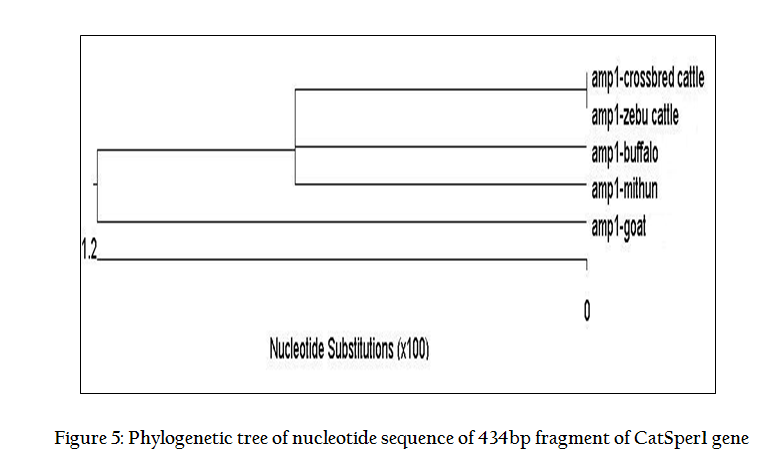
Digestion of 434 bp amplicon corresponding to exon 2, with EcoRI restriction enzyme yielded two bands of 315 bp and 119 bp (Figure 1) in all the animals under study. This restriction pattern was due to the presence of only one restriction site (G↓AATTC) at the 315th nucleotide of amplicon. Similarly, HindIII (A↓AGCTT) having a restriction site at 74th position yielded two bands of 360 bp and 74 bp (Figure 2). These type of bands suggested that this amplicon contained only one RE site for both the restriction enzymes. All the animals under study were found to be monomorphic with reference to these restriction enzymes. The genotype was deduced to be of homozygous normal type in all individuals. Contrary to this, Modi et. al. (2011) observed polymorphism in exon 2 by using PCR–SSCP technique and revealed two allelic patterns in Vrindavani as well as Tharparkar cattle. Monomorphic pattern has also been observed in exon 5 of CatSper1 gene in Vrindavani cattle using restriction enzymes AluI and TaqI (Geetha et al., 2011). However, polymorphism was observed in this gene by using PCR–SSCP method by Modi et al. (2011) in these regions too. Sivakumar et al (2013a and 2013b) observed various single nucleotide polymorphisms (SNPs) in CatSper1 and CatSper2 genes in Vrindavani as well as Tharparkar cattle. The sequence of 434 bp amplicon of CatSper1 gene of crossbred cattle (GU372964) was compared with similar sequences of other ruminants viz zebu cattle (GU372965), mithun (GU372966), buffalo (GU372967) and goat (GU372968). On comparison of crossbred cattle with zebu cattle, no nucleotide variation was seen; however, one nucleotide variation was observed in mithun in the intron 1 and four in the exon 2. Six nucleotide differences were noticed, when crossbred cattle sequences were compared to buffalo; all of them were found in exon 2. Similarly, when nucleotide sequence of crossbred cattle was compared with goat, one nucleotide variation was found in intron1, whereas, eleven nucleotide variations were present in exon 2. Only buffalo and goat had four amino acid changes when compared to crossbred cattle. The nucleotide sequence homology of crossbred cattle was found to be highest with zebu cattle (100%). The deduced amino acid sequences showed 100% homology with zebu cattle and mithun.
A fairly good number of crossbred bulls are known to donate semen of poor quality leading to high rejection rate of the bulls in breeding programmes (Mathew et al., 1982; Rao and Rao, 1991 and Kumar, 2006). Genetic characterization and polymorphism identification of CatSper1 is prerequisite for finding out a genetic marker of this gene which may help in improvement of sperm motility and freezability in crossbred cattle through marker assisted selection. The monomorphic pattern of exon 2 of CatSper1 gene with respect to different enzymes suggested to explore polymorphism using other techniques viz. PCR–SSCP and also in other coding regions of this gene. Work is also required to explore other genes responsible for sperm motility. Further, if polymorphism in this gene is identified in future, then association of these polymorphs with sperm motility could be evaluated, which would be another step in developing a genetic marker for selection programme.
REFERENCES
Chacon J, Perez E, Muller E, Soderquist L and Martinez HR (1999). Breeding soundness evaluation of extensively managed bulls in Costa Rica. Therio. 52: 221– 231.
http://dx.doi.org/10.1016/S0093-691X(99)00124-7
Darszon A, Acevedo JJ, Galindo BE, Hernandez–Gonzalez EO, Nishigaki T, Trevino CL, Wood C and Beltran C (2006). Sperm channel diversity and functional multiplicity. Reprod. 131: 977– 998.
http://dx.doi.org/10.1530/rep.1.00612
PMid:16735537
Dhanju CK, Ranjha S, Cheema C and Kaur P (2006).Correlation of Animal spermatozoalfreezability with semen characteristics in crossbred bulls. Ind. J. Anim. Sci.76 (3):241– 243.
Geetha T, Kumar S, Dubey PP, Sivamani B, Ghosh SK, Mitra A, Tomar AKS and Sharma A. (2011). Sequence variability in CatSper1 gene in Vrindavani crossbred cattle.Ind. J. Anim. Sci.81 (9): 981– 983.
Ghosh SK, Singh SK, Singh LP, Tripathi RP and Tumnyak L (2007).Rejection rate in crossbred bull semen.Compendium XXIII Annual Convention and National Symposium on Challenges in improving reproductive efficiency of farm andpet animals. Bhubaneshwar, Orissa, December 7– 9.
Jin JL, O'DohertyAM, Wang S, Zheng H, Sanders KM and Yan W (2005). CatSper3 and CatSper4 encode two cation channel–like proteins exclusively expressed in the testis. Biol. Reprod. 73: 1235– 1242.
http://dx.doi.org/10.1095/biolreprod.105.045468
PMid:16107607
Kumar S (2006). Advances in assessment of frozen semen of crossbred bulls. National Seminar on Artificial Insemination: Acceptability, impact, constraints and solutions. New Delhi, India, September 21–14, 2006. pp. 106– 116.
Lobley A, Pierron V, Reynolds L, Allen L and Michalovich D (2003). Identification of human and mouse Catsper3 and Catsper4 genes: characterization of a common interaction domain and evidence for expression in testis. Reprod. Biol. Endocrinol.1: 53 – 67.
http://dx.doi.org/10.1186/1477-7827-1-53
PMid:12932298 PMCid:PMC184451
Mathew A, Joseph PJ, Jose TK (1982). Semen characteristics of purebred and cross–bred bulls. Ind. Vet. J. 59: 364– 367.
Modi RP, Kumar S, Dubey PP, Sivakumar A, Kumar A, Sivamani B, Ghosh SK, Mitra A and Sharma A (2011). Genetic variability in cation channel of sperm (CatSper1) gene in cattle. National Symposium on Reproductive biotechnologies for augmenting fertility & conservation of animal species with special reference to NEH Region, Aizawl, Mizoram, India, 29 September 27–29, 2011. p. 111
Qi H, Moran MM, Navarro B, Chong JA, Krapivinsky G, Krapivinsky L, Kirichok Y, Ramsey IS, Quill TA and Clapham DE (2007). All four CatSper ion channel proteins are required for male fertility and sperm cell hyperactivated motility. Proc. Natl.Acad. Sci. U. S. A., 104: 1219– 1223.
http://dx.doi.org/10.1073/pnas.0610286104
PMid:17227845 PMCid:PMC1770895
Quill TA, Ren D, ClaphamDE andGarbers DL (2001). A voltage–gated ion channel expressed specifically in spermatozoa.Proc. Natl. Acad. Sci. U. S. A. 98: 12527– 12531.
http://dx.doi.org/10.1073/pnas.221454998
PMid:11675491 PMCid:PMC60087
Rao BK and Rao AR (1991).Evaluation of crossbred bull for breeding soundness.Ind. J. Anim. Reprod. 12: 111–113.
Ravimurugan T, Kanakaraj P and Thangaraju P (2007).Frozen semen production potential of murrah bulls.Tamilnadu J. Vet. Anim. Sci. 3(5): 269– 271.
Ren D, Navarro B, Perez G, Jackson AC, Hsu S, Shi Q, Tilly JL and Clapham DE (2001). A sperm ion channel required for sperm motility and male fertility. Nature., 413: 603– 609.
http://dx.doi.org/10.1038/35098027
PMid:11595941
Sambrook J and Russel DW (2001).Molecular cloning– A laboratory manual.3rd edition.Cold Spring Harbor Laboratory Press. New York.
Sivakumar A, Kumar S, Dubey PP, Yathish HM and Sivamani B (2013a).SNP identification in cation channel of sperm2 (CatSper2) gene in cattle. In National Symposium on "Integrated Development of Vast Biodiversity of Indigenous Livestock for Long Term Rural Livelihood Security" held art o be held at the Department of Animal Genetics and Breeding, College of Veterinary and Animal Sciences, Pantnagar during 7th–8th February, 2013.
Sivakumar A, Kumar S, Dubey PP, Yathish HM and Sivamani B (2013b). Variability in cation channel of sperm1 (CatSper1) gene in cattle. InNational Seminar on Technological and Policy Interventions of Sustainable Cattle Breeding in India at Project Directorate on Cattle, Meerut on 14th Mar 2013
Tyagi S, Mandal DK, Kumar M and Mathur AK (2006).Reproductive wastage rate of crossbred dairy bulls with reference to level of exotic inheritance and number of breed component.Ind. J. Anim. Reprod. 27(1): 27– 30.

