
Advances in Animal and Veterinary Sciences


Research Article
Immunoreactivity to Culture Filtrate Proteins of Mycobacterium avium Subspecies paratuberculosis in Naturally Infected Goat and Sheep Sera
Saurabh Gupta1, 2, Kundan Kumar Chaubey1, Shoor Vir Singh1*, Ashok Kumar Bhatia2, Naveen Kumar1, Anjana Goel2, Tarun Kumar Sachan1, Krishan Dutta Rawat1, Jagdip Singh Sohal3, Kuldeep Dhama4
1Microbiology Laboratory, Animal Health Division, Central Institute for Research on Goats, Makhdoom, PO-Farah, Mathura, Uttar Pradesh, India; 2Department of Biotechnology, GLA University, Chaumuhan, Mathura, Uttar Pradesh, India; 3Amity Institute of Microbial Technology, Amity University Rajasthan, Jaipur, India; 4Division of Pathology, Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh, India.
Abstract | Mycobacterium avium subspecies paratuberculosis (MAP), the cause of granulomatous chronic enteritis in ruminants (Johne’s disease) is under reported due to difficulties in diagnosing pre-clinical cases. Compromised specificity is a problem due to extensive sharing of antigens /epitopes among MAP and other mycobacterial strains. Culture filtrate (CF) proteins profile of native ‘Indian Bison Type’ strain of MAP was studied at different harvesting times (2-8 weeks of growth) in Middlebrook 7H9 medium supplemented with ADC, PANTA antibiotics and mycobactin J. Analysis of harvested CF proteins by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) showed that the greater part of CF proteins had molecular masses (<70 kDa) as 14, 19, 26, 34-41, 52-55, and 70 kDa. Immunoblotting showed reactivity of CF proteins commonly recognised (28, 34-36, 38-42, 45, and 56 kDa) with all four MAP infected goat and sheep sera at 2-8 weeks of growth. Collectively, these immunoreactive MAP CF proteins could be the potential targets for developing diagnostics against Johne’s disease with improved sensitivity and high specificity instead of whole cell sonicated crude protoplasmic extracts (PPA).
Keywords | Johne’s disease, Mycobacterium avium subspecies paratuberculosis, Culture filtrate, SDS-PAGE, Immunoblotting
Editor | Muhammad Zubair Shabbir, Assistant Professor, Quality Operations Laboratory, University of Veterinary and Animal Sciences, Lahore, Pakistan.
Received | March 28, 2015; Revised | April 23, 2015; Accepted | April 24, 2015; Published | May 17, 2015
*Correspondence | Shoor Vir Singh, Central Institute for Research on Goats, Makhdoom, Mathura, Uttar Pradesh, India; Email: shoorvir_singh@rediffmail.com
Citation | Gupta S, Chaubey KK, Singh SV, Bhatia AK, Kumar N, Goel A, Sachan TK, Rawat KD, Sohal JS, Dhama K (2015). Immunoreactivity to culture filtrate proteins of Mycobacterium avium subspecies paratuberculosis in naturally infected goat and sheep sera. Adv. Anim. Vet. Sci. 3(6): 347-353.
DOI | http://dx.doi.org/10.14737/journal.aavs/2015/3.6.347.353
ISSN (Online) | 2307-8316; ISSN (Print) | 2309-3331
Copyright © 2015 Gupta et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Introduction
Mycobacterium avium subspecies paratuberculosis (MAP), a cause of Johne’s disease (Paratuberculosis) is the most wide-spread and highly prevalent due to difficulties in diagnosing pre-clinical cases. MAP has wide host range including domestic and wild ruminants, free grazing animals and also the human beings (Singh et al., 2010; Singh et al., 2014a; Singh et al., 2014b). Paratuberculosis has high economic impact on dairy industry and > USD 250 million economic losses has been reported in US alone (Ott et al., 1999). Though disease is endemic in India and economic losses in dairy farm was estimated from reproductive disorders (Rs. 23400.0/cow/year), forced removal (Rs. 41,750.0/cow/year), reduced milk yield (Rs. 5,712.0/cow/ year) and increased mortality (Rs. 11,666.0/cow/year) (Rawat et al., 2014). Control of paratuberculosis has been hindered due to lack of efficient and accurate diagnostic tests. Lower specificity of tests can also be a problem due to the sharing of antigens or epitopes among MAP and other mycobacteria (Collins et al., 1991). Sensitivity of commercially available ELISA kits prepared using protoplasmic antigen has been reported comparatively low, 13.6-33.3% (Singh et al., 2007).
Recent studies have focused on developing improved serodiagnostics using species-specific multiple protein antigens (Singh et al., 2014c). Microfluidics and Lab-on-Chip are some of the recent technologies that can predicate development of laboratory-free diagnostic devices for mycobacterial infections (Wadhwa et al., 2012a; Li et al., 2011). Earlier reports have identified several antigens inducing strong antibody responses but most of them have been found unsuitable for serodiagnosis because they are highly conserved within mycobacteria and hence cross-react with other mycobacterial pathogens. Recently, various MAP specific proteins/genes has been characterized, cloned, expressed and the protein was evaluated for their diagnostic potential (Cho et al., 2007). However, strain of MAP may also tune the sensitivity and specificity of test. Present study investigated CF protein profile of native Indian bison type (‘S 5’) MAP strain for the first time with reference to determine the antigenicity / reactivity of the CF proteins by immunoblotting with polyclonal antisera of goat and sheep infected with MAP and to develop diagnostic tests of higher sensitivity and specificity.
Materials and methods
Mycobacterial Strain
Mycobacterium avium subspecies paratuberculosis (‘S 5’) strain was procured from the mycobacterial repository, Central Institute for Research on Goats (CIRG), Makhdoom. The strain was maintained on Modified Herrold’s egg yolk medium with mycobactin J (HEYM) as per Singh et al. (1996) and was sub-cultured in Middlebrook 7H9 medium (as per the manufacturer, Becton Dickinson, BD) supplemented with ADC (10% or 100ml/l), PANTA antibiotics and mycobactin J (2mg/l).
Growth Pattern
Growth of MAP in middlebrook 7H9 medium was monitored by taking absorbance (OD) of the culture at third day, then every week at 600nm for a period of 8 weeks (Figure 1). Growth curves were obtained by plotting the absorbance (OD) values versus incubation time in weeks. Turbidity in the medium was measured by Mcfarland standards (0.5, 1, 2, 3, 4, 5 and 6) to check the growth of the culture. To avoid clump formation the cultures were constantly shaken at 100 rpm during incubation.
Culture Filtrate (CF)
CF were obtained by harvesting (4000×g, 20 min, 4°C) bacterial growth at different times points (2, 4, 6 and 8 weeks of incubation) and was filtered (using 0.22-μm pore size syringe filter, Millipore). CF were precipitated with saturated ammonium sulfate (Rankem) due to the high amount of albumin in the broth followed by extensive dialysis overnight against 10 mM phosphate-buffered saline (pH 7.4) until free of ammonium ions at 4°C. Dialysed CF were concentrated 10-fold using vacuum concentrator (SPD Speedvac, Thermo Savant). Concentration of CF proteins was quantified by Bradford protein assay kit (Genei) and was stored at -20°C.
Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis (SDS- PAGE)
CF protein profile was analyzed by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) under reducing conditions and stained with Coomassie brilliant blue as per method given by Laemmli (1970).
Immunoblotting
The electrophoresed CF proteins were transferred on to PVDF-plus membrane (pore size 0.45 µm, Millipore) using Mini Trans-blot cell (Bio-Rad) for 1.5 hrs at 60 V, 100 mA as per Towbin et al. (1979) with some modifications. Membrane was blocked for 1 hr at room temperature in 5% skim milk (Himedia) containing TBS with 0.05% Tween-20 (TBST) and were probed with polyclonal primary antibody (goat serum) diluted 1:100 in 1X TBST for 2 hrs at room temperature. After incubation membrane was washed 3 times with 1X TBST. Reactivity was seen by incubating the PVDF-plus membrane with peroxidase-conjugated anti-goat IgG (Sigma, USA) in 1:2000 dilution for 1 hr at room temperature followed by washing 5 times (5 min each) with TBST. Visualization of immuno-reactive protein bands was done by diaminobenzidine (DAB) (Sigma).
Statistical Analysis
The correlation (R2) and Standard deviation of OD600 with time interval of growth measurements were analyzed using Graph Pad InStat.
Results
Growth Pattern
Mycobacterium avium subspecies paratuberculosis (‘S 5’) strain developed a granular growth with visible clumps suspended in the 7H9 medium up to 6 weeks, afterwards, clusters of floating cells were observed (Figure 1). The growth curve of MAP ‘S 5’ strain in 7H9 medium showed sharp bacterial growth phases delineation; however, after 3 weeks of incubation the culture was in log phase (early exponential) upto 8 weeks (mid to late exponential) in 7H9 medium (Figure 1). Total bacterial mass was with a maximum absorbance of 1.43 at 8 weeks. OD600 and time interval of growth measurements were highly correlated (R2 = 0.997, Figure 1). We determined in Indian Bison type ‘S 5’ strain of MAP that 1 McFarland unit was nearly equivalent to 0.26 OD600.
Table 1: History of goats and sheep whose serum were used in the immunoblotting
|
Sn |
Animal type/Breed |
Age/ Sex |
Physical condition |
Diarrhea |
Fecal Shedding (Microscopy) |
IS900 blood PCR |
Serology (‘Indigenous ELISA’) |
|
1 |
Sheep (Muzzaffarnagri) |
Adult female |
Extremely weak and emaciated |
Yes |
Yes |
Positive |
Positive |
|
2 |
Goat 1 (Barbari) |
Adult female |
Weak and emaciated |
Yes |
Yes |
Positive |
Positive |
|
3 |
Goat 2 (Jamunapari) |
Adult female |
Weak and emaciated |
No |
Yes |
Positive |
Positive |
|
4 |
Goat 3 (Jakhrana) |
Adult male |
Weak |
Yes |
Yes |
Negative |
Positive |
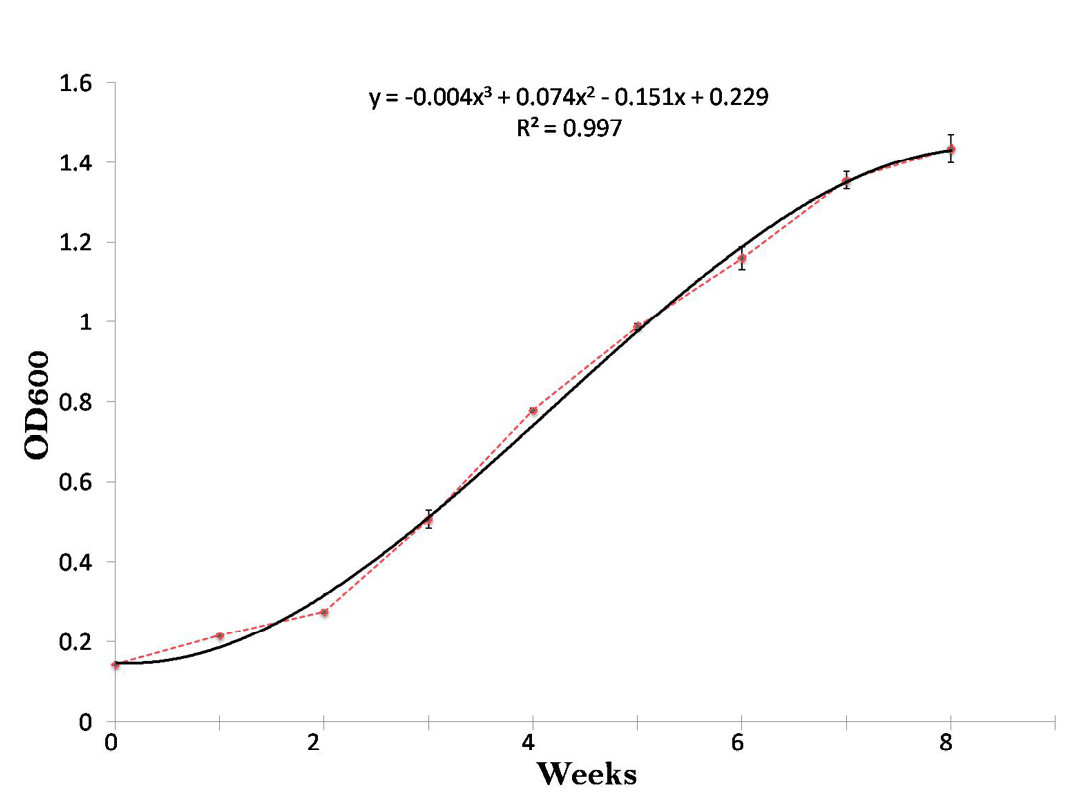
Figure 1: Growth curve of Mycobacterium avium subspecies paratuberculosis (‘S 5’) strain upto 8 weeks by taking OD at 600nm. Each point correlates to the mean of three determinations ± standard deviation
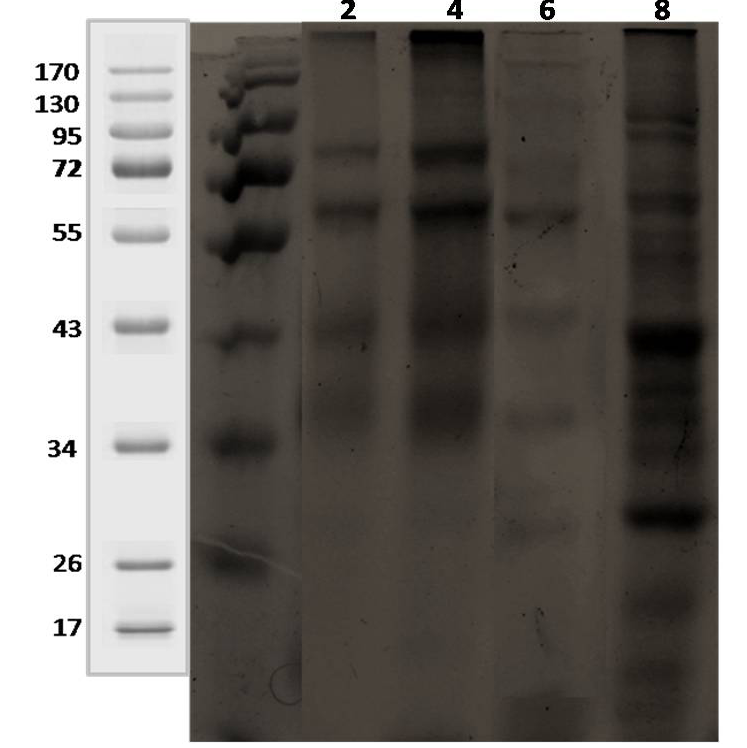
Lane 1: 2 weeks; lane 2: 4 weeks; lane 3: 6 weeks; lane 4: 8 weeks.
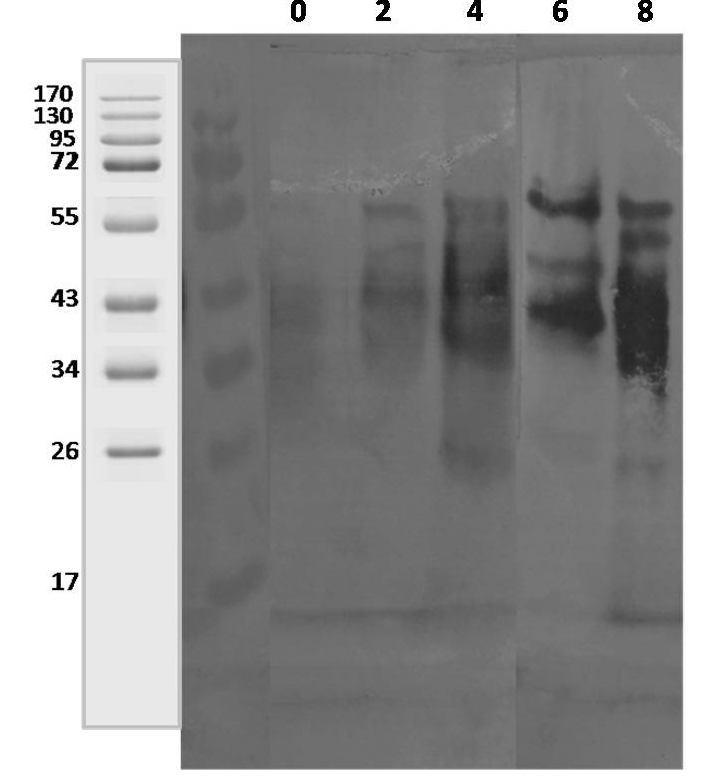
Lane 1: 0 Day; lane 2: 2 weeks; lane 3: 4 weeks; lane 4: 6 weeks; lane 5: 8 weeks.
Immunoblot of Mycobacterium avium Subspecies paratuberculosis Secreted Proteins
The initial antigenic profiles were detected at 2, 4, 6 and 8 weeks of incubation in 7H9 medium. Analysis of harvested CF proteins by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) showed that the greater part of CF proteins had molecular masses (<70 kDa) as 14, 19, 26, 34-41, 52-55, 68-70 and 90-92 kDa (Figure 2). The antibody recognition patterns of secreted proteins using 4 positive sera of goats and sheep (Table 1) showed a high degree of variability among MAP infected animals (heavy shedders). Multiple proteins were bound by all the four positive sera, but the specific proteins bound by each serum sample were variable. However, the antigenic CF proteins of MAP ‘S 5’ strain which showed wide variation in recognition pattern with sera from clinical cases were ranged from 19 to 62 kDa at 2 to 8 weeks of growth (Figure 3, 4 and 5).
Antibody Recognition Pattern to Secreted proteins using sera from 4 Clinical Cases
Polyclonal sera from 3 goats and 1 sheep naturally infected with Mycobacterium avium subspecies paratuberculosis (confirmed by fecal microscopy, IS900 blood PCR and ‘Indigenous serum ELISA kit’) were tested for reactivity pattern to CF proteins. The CF proteins commonly recognised with all four MAP infected goats and sheep sera at 2 weeks (early growth) were approximately, 38-42, 45, and 56 kDa and at 8 weeks growth were approximately, 28, 34-36, 38-42, 45, and 56 kDa. Sera from goat 1, 2 and 3 reacted with 28, 38, 45 and 65 kDa secreted proteins at 2-8 weeks of growth (Figure 4 and 5). However, 38-42, 45 and 56 kDa CF proteins reacted with sheep serum (Figure 3).
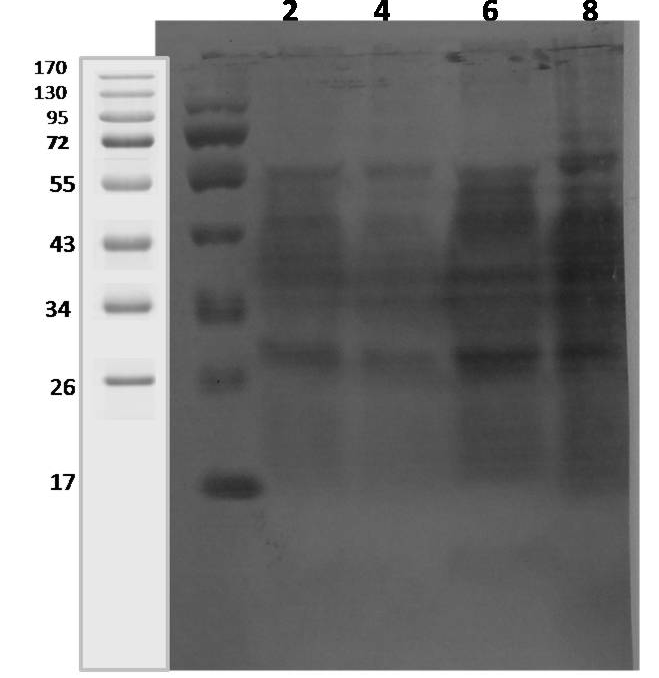
Lane 1: 2 weeks; lane 2: 4 weeks; lane 3: 6 weeks; lane 4: 8 weeks.
Discussion
Current immuno-diagnostics for Johne’s disease, a chronic enteritis in ruminants, that is responsible for extensive economic losses to farmers and dairy industry worldwide caused by Mycobacterium avium subspecies paratuberculosis (MAP) are hampered by the lack of specific antigens. The capability of rapidly diagnosing the disease and identifying its causative agent is critical to combat diseases and halt epidemics (Li et al., 2014). Test results from the sub-clinically MAP infected animals are challenge to interpret, because clinical signs are not present to assist the interpretation. Transmission of infection usually takes place prior the tests becoming positive and before clinical signs developed. Most frequently used test for diagnosis of MAP infection include bacterial culture, IS900 PCR amplification, interferon-γ assay and serum antibody detection using an ELISA platform. Wadhwa et al. (2012) used complement fixation, agar gel immunodiffusion (AGID), and ELISA to determine the antibody response in JD. The complement fixation and agar gel immunodiffusion tests both suffer poor sensitivity (Sherman et al., 1990; Pahangchopi et al., 2014), series of studies reported that immunological tests like ELISA is the best method as compared to complement fixation and AGID for controlling JD in dairy and beef herds (Wynne et al., 2011). Recent studies have focused to develop improved serodiagnostics using species-specific multiple protein antigens. Detection of MAP using ELISA have been reported in many previous studies using different antigens as protoplasmic antigen (PPA) (Collins et al., 2005; Singh et al., 2007; Sharma et al., 2008), lipoarabinomannan (LAM) (Sweeney et al., 1994), culture filtrate of MAP (Shin et al., 2008), and MAP proteins (Map1152, Map1156, Map2609, Map2942c and Map0210c) (Bannatine et al., 2011; Willemsen et al., 2006) for testing antibodies against MAP. Although PPA contains proteins very similar to proteins commonly found in closely related mycobacteria species. LAM is one of the constituent of the cell wall of mycobacteria species and its core structure is shared among mycobacterial species (Mishra et al., 2011). Some studies also suggested that secreted proteins may be better as solid-phase ELISA antigens resulting in a more sensitive assay (Cho and Collins, 2006; Willemsen et al., 2006). On the other hand, Sung and Collins (2003) have reported that expression of MAP CF protein depends on the type and culture conditions.
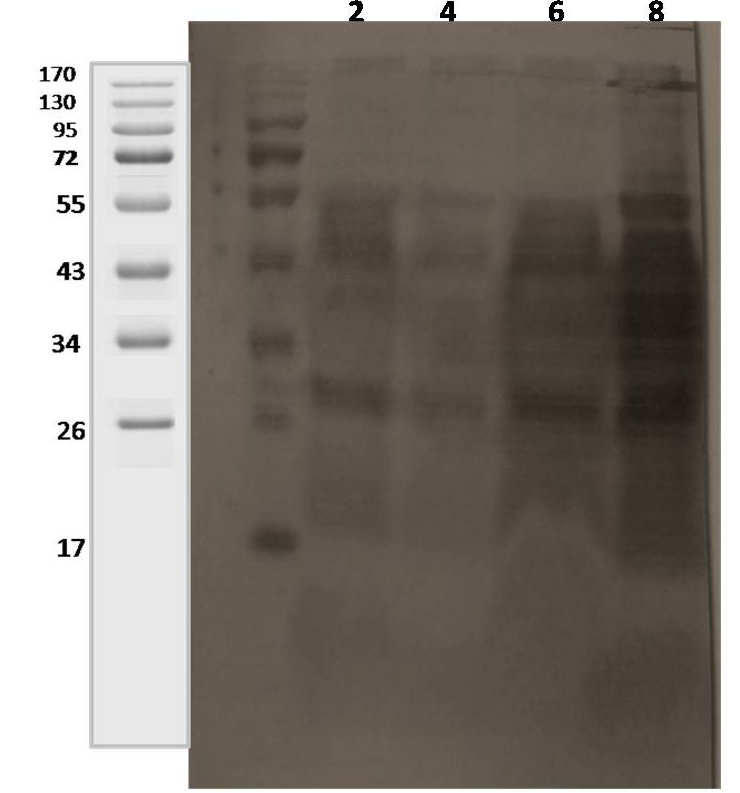
Lane 1: 2 weeks; lane 2: 4 weeks; lane 3: 6 weeks; lane 4: 8 weeks.
Recent technological developments have led to the proliferation of new, rapid diagnostic tests that hold promise for the improved management and control of infectious diseases. Mycobacterial infections such as tuberculosis (TB), bovine tuberculosis (bTB), and Johne’s disease (JD) are major infectious diseases of both human and animals (Wadhwa et al., 2012b). For tuberculosis and bovine TB, there has been recent progress in developing laboratory-free diagnostic methods. New technologies such as ‘microfluidics’ (Wadhwa et al., 2012a) and ‘Lab-on- Chip’ (Li et al., 2011) are examples of promising new technologies that can underpin development of laboratory-free diagnostic devices for these mycobacterial infections. Although there have been developed a sensitive serum ELISA test, ethanol vortex enzyme-linked immunosorbent assay (EVELISA), using ethanol extract of MAP especially for Johne’s disease (Wadhwa et al., 2012c). Another study was conducted to assess the performance of EVELISA optimized to diagnose bovine TB using serum samples from various groups of red deer and cattle including animals experimentally infected with MAP and suggests that EVELISA can form a basis for development of a sensitive and specific test for bovine TB (Wadhwa et al., 2013; Wadhwa et al., 2014). Furthermore, EVELISA-based control measures increase the annual per capita revenue of US dairy farms when compared to no JD control and ELISA based JD control, respectively (Massaro et al., 2013). An AC electrokinetic impedance sensing can be used for rapid and sensitive detection of specific antibodies in serum samples and this could be a basis for development of a point of care diagnostic device for human and bovine tuberculosis (Cui et al., 2013). Garg et al. (2015) recently reported that the combination of milk ELISA and milk PCR may be adopted as a model strategy for the screening and diagnosis of JD in lactating cattle herds in Indian conditions.
Our results demonstrate the correlation between turbidity and OD600 measurements with time period of growth. We determined in Indian Bison type ‘S 5’ strain of MAP that 1 McFarland unit was nearly equivalent to 0.26 OD600. These measurements were not quite as precise, since Mycobacterium avium subsp. paratuberculosis tends to form clumps. Strain to strain variations were reported in growth curves and growth rate of MAP (Elguezabal et al., 2011). The present study also showed that 7H9 medium allowed to obtaining more and earlier CF proteins and nearly all these CF proteins were of low molecular weight ranging between 14 and 70 kDa; similar findings regarding CF proteins of MAP have been reported with other field strains elsewhere (Olsen et al., 2000; Cho and Collins, 2006). When immunoblotting test using CF proteins of MAP were performed, a high variability among naturally infected animals in protein binding was observed, similar results were reported by Cho and Collins (2006). Waters et al. (2004) have shown variable antibody binding patterns in assays with serum from white-tailed deer experimentally infected with M. bovis. High correlation has been shown between the level of shedding of MAP in feces and concentration of serum antibodies (Nielsen and Toft, 2006). However, in agreement, our results showed high intensity and more number of bands were recognized with naturally infected goats and sheep sera. Several other factors may also affect serum antibody levels (Hendrick et al., 2006), including individual genetic variation (Sudgen et al., 1997). ValentinWeigand and Moriarty (1992) also reported that during short incubation period MAP may secrete immunoreactive proteins which are not dominant in long term cultures.
There are studies evaluating sensitivity and specificity of commercial ELISA kits in domestic and wild ruminants derived from bovine strains of MAP (Whittington et al., 2003) however, ‘Indigenous ELISA’ developed from ‘Indian Bison type’ strain showed improved sensitivity and specificity as compared to commercial ELISA kits (Singh et al., 2007). Conceptually, the use of specific purified proteins as the solid phase antigen in ELISA assay have suffered from low diagnostic sensitivity (Huntley et al., 2005). Recombinant antigens can also alter antigenicity due to conformational changes or lack of post-translational modifications (Cho et al., 2007; Li et al., 2007). The use of MAP CF proteins as antigen in ELISA assay increased sensitivity by 25% over commercial ELISA kits for low shedding animals (Shin et al., 2008). Also the variability in responses among different animals to recognize single MAP-specific antigen in early and subclinical stages, a cocktail of immuno-reactive native proteins of ‘Indian Bison type’ provide significant advantages for improved serodiagnosis of MAP infection. Moreover, the immunoblotting findings concluded that the CF proteins were quite specific and the immunogenecity of these reactive CF proteins would be further evaluated as biomarkers to develop more sensitive and specific assays.
Acknowledgement
Authors are thankful to Directors (Central Institute for Research on Goats, Makhdoom, Farah and GLA University, Mathura) for providing laboratory and aanimal house facilities.
Conflict of Interest
No potential conflict of interest to declare.
ReferenceS

