
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 1 (4S): 14 – 19Special Issue–4 (Progress in Research on Viruses and Viral Diseases)
Evidence of Reassortment between Eastern and Western Topotype Strains of Bluetongue Virus Serotype 16 (BTV–16) from India
Pawan Kumar, Prasad Minakshi1*, Koushlesh Ranjan1, Ravinder Dalal1, Gaya Prasad1,2
- Department of Animal Biotechnology, College of Veterinary Sciences, Lala Lajpat Rai University of Veterinary and Animal Sciences, Hisar 125004, Haryana, India
- Indian Council of Agricultural Research, Krishi Bhawan, New Delhi 110001, India
*Corresponding author:minakshi.abt@gmail.com
ARTICLE CITATION:
Kumar P, Minakshi P, Ranjan K, Dalal R and Prasad G (2013). Evidence of reassortment between eastern and western topotype strains of bluetongue virus serotype 16 (BTV–16) from India. Adv. Anim. Vet. Sci. 1 (4S): 14 – 19.
Received: 2013–10–18, Revised: 2013–11–10, Accepted: 2013–11–10
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/134/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
The dsRNA genome of bluetongue virus (BTV) consists of 10 segments that encodes for seven structural and four non–structural proteins. The nucleotide variations of core, NS1 and NS2 proteins, in addition to capsid proteins helps determine the geographical origin (eastern or western geographical group/ topotype) of virus isolates. International trade of vaccinated livestock are playing major role in intercontinental movement of western BTV strains or genome segments in India. The co–circulation of different virus isolates, being the segmented nature of genome, provides an opportunity to simultaneously infect the same animal or even same cell with the potential of generating reassortant viruses. The data regarding the prevalence of viral topotype/ nucleotype/ serotype is essential to establish phylogenetic relationship and intercontinental migration of the pathogen which in turn help in formulation of proper intervention strategies against the disease. We carried out full length sequencing of genome seg–5 (ns1 gene) of Indian isolate VJW64 (BTV16). Sequence identity and phylogenetic analysis revealed that genome seg–5 clusters with isolates of western topotypes showing nucleotide similarity of more than 89.5% compared to the less than 78% sequence identity with isolates of eastern geographical group that formed a separate monophyletic clade. However our previous study indicated that seg–2 and seg–6 of Indian isolate VJW64 (BTV16) belong to eastern topotypes of bluetongue virus. Thus VJW64 isolate is a reassortant virus containing segments derived from both eastern and western topotypes. The study provides an evidence of circulation of a reassortant of BTV16 serotype in India for the first time.
INTRODUCTION
The bluetongue virus (BTV) causes an economically important arthropod borne (Culicoides spp.) viral disease of ruminants. BTV is a prototype member of genus Orbivirus, belonging to family Reoviridae. The genome of BTV consists of 10 segments of double stranded RNA located in core, which encodes for seven structural (VP1 to VP7) and four non–structural proteins (NS1, NS2, NS3/3A and NS4) (Belhouchet et al., 2011). The core of virus which composed of two major (VP3 and VP7) and three minor (VP1, VP4 and VP6) proteins is further surrounded by outer capsid consisting of two major proteins, VP2 and VP5. The outer capsid proteins are highly variable and responsible for virus neutralization and serotype determination (Huismans et al., 1987; Mertens et al., 1989; Cowley and Gorman, 1989). The core proteins, NS1 and NS2 are highly conserved and antigenically cross reactive between different BTV serotypes. Although, these conserved proteins show no significant correlation with virus serotype, they do show sequence variations that reflect the geographical origin (topotype) of virus isolate (Gould and Pritchard, 1990; Balasuriya et al., 2008; Nomikou et al., 2009).
A total of 26 BTV serotypes (Prasad, 2000; Hofmann et al., 2008; Maan et al., 2011) and two major geographical groups (eastern and western topotypes) (Balasuriya et al., 2008; Nomikou et al., 2009) of BTV strains have been identified worldwide.
Gene mutation, segment reassortment and intragenic (homologous) recombination are key forces that drive evolution of BTV and led to emergence of novel BTV lineages, strains and serotypes (He et al., 2010; Shaw et al., 2013). The segmented nature of BTV genome allows the reassortment of BTV segments when host cell is concurrently infected by two or more different viral strains or serotypes (Gorman, 1990; Batten et al., 2008). This led to the development of viral diversity (Carpi et al., 2010) and changes in virulence and serological characteristic of the virus (Mertens et al., 1989).
Live attenuated vaccines have been used to control the disease in USA, South Africa and Mediterranean region. These vaccine viruses are associated with low level of viraemia and can spread to vectors with potential for reversion to virulence and/or reassortment with genes of wild type virus (Veronesi et al., 2005; Savini et al., 2009). Live attenuated vaccine viruses and trade of livestock and their products are playing major role in intercontinental movement of western BTV genome segments in India (Rao et al., 2012). Till date, reassortant strains of eastern and western topotypes of BTV2 (Maan et al., 2012c), BTV3 (Maan et al., 2012a) and BTV23 (Maan et al., 2012e) have been reported from India. All of these isolates were reported to have seg–5 of western origin and other segments of eastern lineage. A reassortant Indian isolate of BTV21 (KMNO–7) with segment 6 (seg–6) from BTV16 isolate (VJW64) have been reported by Shafiq et al. (2013).
The knowledge regarding the prevalence of different serotypes and topotypes is prerequisite for successful vaccination strategies and epidemiological studies. To know the geographical origin of seg–5 of BTV16 isolate (VJW64), we carried out full length amplification and sequencing. The nucleotide sequence was analyzed and phylogenetic relationship with various global isolates of BTV was determined.
MATERIALS AND METHODS
Virus Isolate
The BTV16 isolate (designated as VJW–64/08/IND) was obtained in an outbreak from Andhra Pradesh (India) and was isolated by inoculating blood samples of clinically affected sheep in embryonated chicken eggs.
Virus Cultivation in Cell Culture
Baby Hamster kidney–21 (BHK–21) (clone 13) cell line, originally obtained from National Cell Science Centre, Pune, India was used for virus propagation as described by Jain et al. (1986). The cell cultures which attained 70–80% confluency were inoculated with virus isolate. One ml of virus suspension was used per 75–cm2 tissue culture flask (Greiner Biocon).
Extraction of Viral Genomic RNA
The cell sheet was detached from tissue culture flask in the growth medium and centrifuged in 15 ml plastic tubes at 2000 rpm (RCF– 750 x g) (VELOCITY 18R, Dynamica) for 15 min. The pellet was resuspended in 0.5 ml of supernatant and used for RNA extraction by TRIZOL (Life Technologies) method as per manufacturer’s protocol.
RNA–Polyacrylamide Gel Electrophoresis (RNA–PAGE)
After extraction, the segmented RNA of virus was analyzed by RNA–PAGE using discontinuous buffer system without SDS as described by Laemmli (1970). The gel was stained with silver nitrate as described by Svensson et al. (1986).
Reverse Transcription–Polymerase Chain Reaction (RT–PCR)
The viral genomic dsRNA was reverse transcribed using random primers (decamer) (Ambion) and moloney murine leukemia virus reverse transcriptase (Mo–MuLV–RT) enzyme (Promega). The resultant cDNA was used as template for PCR amplification of ns1 gene of the virus.
Designing of BTV Genome seg–5 (ns1 gene) Specific Primers
Genome seg–5 specific primers were designed using the BTV sequence (Accession no. FJ713348.1) available in GenBank (http://www.ncbi.nlm.nih.gov/GenBank/) with the help of Pick Primer software of NCBI (Table 1). Primer pair I was designed for full length gene amplification while both primer pairs were used in sequencing PCR.

Amplification of Genome Seg–5 (ns1 gene) of VJW–64 (BTV16) Isolate
The seg–5 of VJW64 isolate was PCR amplified using designed primer pair I. During standardization, final concentration of 0.4 µM primer (both forward and reverse), 200 µM dNTP, 3% DMSO and 0.6 unit of Phusion DNA polymerase (NEB) per 25 µl of reaction mixture was used. Initial denaturation was carried out at 98oC for 90 s followed by three step cyclic denaturation at 98oC for 10 s, annealing at 65oC for 30 s, extension at 72oC for 1 min and final extension at 72oC for 5 min. The PCR amplified product was analyzed by agarose gel electrophoresis (AGE) using 1.0 % agarose (LifeTech) gel containing 0.5 µg ethidium bromide (Sigma) per ml in tris–acetate–EDTA (TAE) buffer along with 1kb DNA ladder (MBI fermentas).
Cloning in pJET1.2 Blunt Vector
The full length PCR amplified product was cloned in pJET1.2 blunt vector supplied with CloneJETTM PCR cloning kit (Fermentas) as per manufacturer’s guidelines. The DH5α strain of Escherichia coli was used as host for transformation with recombinant plasmid vector. The recombinant colonies were screened for the presence of insert of desired length by colony touch PCR using pJET1.2 vector specific primers supplied with kit and by seg–5 specific designed primers. Plasmid isolation was done by alkaline lysis method (miniprep) as described by Sambrook et al. (2001).
Nucleotide sequence Analysis of Genome Seg–5 of VJW–64 Isolate (BTV16)
The recombinant plasmid vector isolated from positive clones was sequenced using ns1 gene segment specific primer pairs (Table 1) and vector specific primers. The nucleotide sequencing was done using automated DNA sequencer ABI PRISMTM 3130 version 3.0 in the Department of Animal Biotechnology, College of Veterinary Sciences, LLRUVAS, Hisar, India. Sequences from different clones were assembled using BioEdit® software (Hall, 1999) to obtain a consensus sequence, which was used for phylogenetic analysis using ClustalX (Larkin et al., 2007), BioEdit® (Hall, 1999) and MEGA 5.0 softwares (Tamura et al., 2011).
RESULTS
The BTV16 isolate (VJW64), originally isolated by inoculating blood samples of clinically affected sheep into chicken embryo, was successfully adapted to BHK21 cell line where it produced BTV specific cytopathic effects (CPE), characterized by cell roundening, detachment, vaculation and clumping of infected cells with bunch–grape like appearance.
The appearance of 10 bands of TRIZOL extracted viral dsRNA genomic segments with typical migration pattern (3:3:3:1) in RNA–PAGE doubly confirmed the nature of infectious agent as BTV. The cDNA synthesized by reverse transcription and amplified by using seg–5 specific primer pair I, yielded an expected 1659 bp PCR product without any non–specific amplification when observed in 1.0% AGE. The pJET vector based screening strategy allows the growth of bacterial colonies transformed only with recombinant vector, as recircularized vector expresses a lethal restriction enzyme. However, further screening of recombinant colonies for presence of ns1 gene insert was performed by colony touch PCR amplification using insert specific (Pair I; Table 1) and vector specific primers that yielded expected size amplicon of 1659 bp and 1778 bp size (Figure 1).
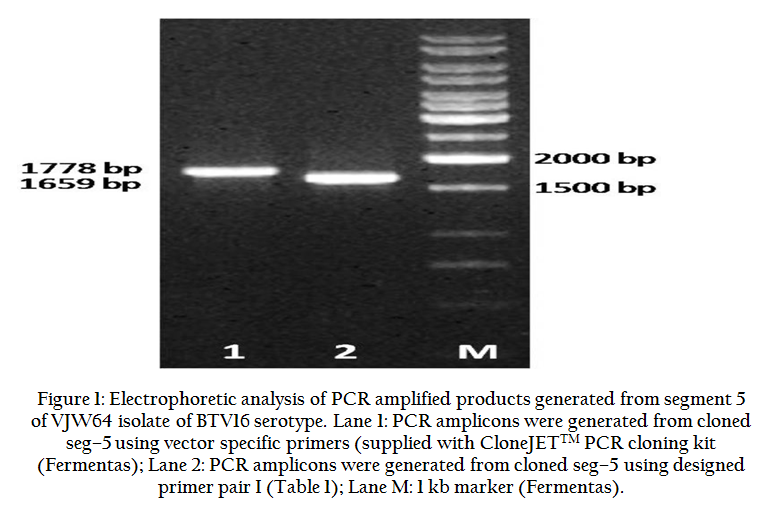
Figure 1: Electrophoretic analysis of PCR amplified products generated from segment 5 of VJW64 isolate of BTV16 serotype
Full length sequence of seg–5 of VJW64 isolate obtained after sequencing as described in materials and methods was submitted to GenBank. Accession number of the sequence generated was KF289016. All the sequences were used to construct sequence identity matrix (Table 2) and neighbor joining phylogenetic tree (Figure 2) and compared thereby.
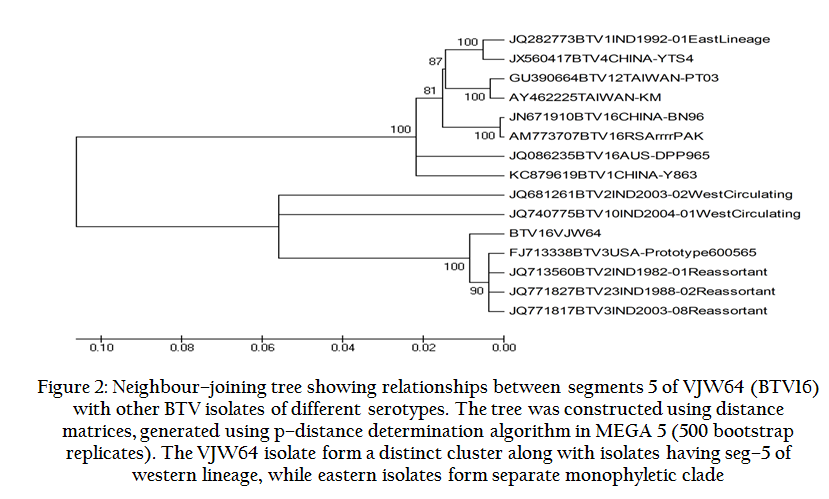
Figure 2: Neighbour–joining tree showing relationships between segments 5 of VJW64 (BTV16) with other BTV isolates of different serotypes.
Genome seg–5 (ns1 gene) of Indian isolate VJW64 (BTV16) belong to major western topotype, showing nucleotide sequence identity of more than 88.5% from global isolates having seg–5 of western origin (Table 2), reflecting western lineage of the segment. These viral strains are either western topotypes [US prototype 600565, Indian BTV2 isolate IND2003/02 (Maan et al., 2012b) and Indian BTV10 isolate IND2004/01 (Maan et al., 2012g)] or they are reassortants of eastern topotype acquiring seg–5 from western topotype [(Indian BTV2 isolate IND1982/01 (Maan et al., 2012c), Indian BTV3 isolate IND2003/08 (Maan et al., 2012a) and Indian BTV23 isolate IND1988/02 ( Maan et al., 2012e)]. Among the isolates of western topotypes, VJW64 isolate was closer to US isolate (prototype 600565) with 98.4% sequence similarity compared to the 88.5–88.6% identity of western isolates [IND2003/02 (Maan et al., 2012b) and IND2004/01 (Maan et al., 2012g)] circulating in India. However, nucleotide sequence identity of genome seg–5 of VJW64 isolate was less than 80% from BTV isolates having seg–5 of eastern lineage (Table 2), including Chinese isolate BN96 (BTV16) and Indian BTV1 isolate IND1992/01 (Maan et al., 2012g).
Table 2: Sequence identity matrix showing nucleotide sequence similarity in genome seg–5 between various global isolates of bluetongue virus (including VJW64 (BTV16)). Nucleotide sequence similarities between VJW64 isolate and global isolates
Further, phylogenetic analysis revealed two major groups (eastern and western topotypes) of various BTV isolates based on seg–5/NS1 gene (Figure 2). BTV isolates having seg–5 of eastern origin were clustered into distinct phylogenetic clade. However, seg–5/ns1 of VJW64 isolate cluster with BTV isolates having seg–5 of western origin forming a separate monophyletic clade, again confirming its western lineage.
The data demonstrates that seg–5/ns1 gene of VJW64 (BTV16) isolate belongs to western topotype of bluetongue virus.
DISCUSSION
Full length amplification and sequencing followed by sequence analysis and phylogenetic relationship are of utmost importance in epidemiological studies. Full genome sequencing of several isolates of different BTV serotypes have been reported so far (Minakshi et al., 2012; Maan et al., 2012a; 2012b; 2012c). Few studies have also revealed the event of reassortment between different isolates (Shafiq et al., 2013).
Due to segmented nature of BTV genome, the virus is highly prone to segment reassortment. This leads to generation of new genetic variants and increased viral diversity. The highly variable capsid (VP2 and VP5) proteins determine the viral serotype and are responsible for type specific serological reaction. The variations of less conserved core, NS1 and NS2 proteins, in addition to capsid protein, are used to identify different topotypes including an eastern group and a western group and thus in determination of the geographical origin of the virus (Gould and Pritchard, 1990; Singh et al., 2004; Maan et al., 2008; Mertens et al., 2007; 2008; Balasuriya et al., 2008; Nomikou et al., 2009).
The appearance of characteristic cytopathic effects of bunch grape like appearance in BHK21 cell culture, presence of 10 discrete genomic segments when observed on RNA–PAGE and full length amplification by ns1 gene specific primers (Figure 1) confirmed the isolate as bluetongue virus.
The sequence identity and phylogenetic analysis presented here shows that genome seg–5/ns1 gene of Indian isolate VJW64 (BTV16) clusters with the global isolates of western topotype and reassortants having seg–5 of western lineage with nucleotide similarity of more than 88.5% (Table 2 and Figure 2). However, isolates of eastern topotype make separate monophyletic clade and show less than 80% sequence identity with VJW64 and other isolates of western group. The data clearly demonstrates the origin of seg–5 of VJW64 isolate from western topotype.
In contrast, earlier study based on nucleotide sequence identity and phylogenetic analysis present that seg–2 (vp2 gene) and seg–6 (vp5 gene) of VJW64 isolate match with the BTV16 isolates of eastern topotype group only, with no similarity to the isolates of western topotype (Shafiq et al., 2013). These isolates include Pakistan isolates RSArrrr, Chinese isolate BN96, several Japanese isolates and other isolates of eastern geographical group. However, seg–5 of these isolates shows very less sequence identity (<80%) with VJW64 and other isolates of western topotype, despite the conserved nature of the segment. This reflects that Indian isolate VJW64 is reassortant strain having seg–2 and seg–6 from eastern topotype and seg–5 from western topotype, providing evidence for the first time the occurrence of reassortant of BTV16 serotype between eastern and western topotype within the Indian subcontinent.
Genetic reassortment on the basis of genome sequencing and phylogenetic analysis has been so far reported for Indian isolates of BTV2 (Maan et al., 2012c), BTV3 (Maan et al., 2012a) and BTV23 serotypes (Maan et al., 2012e).
The co–circulation of western topotype or reassortant of BTV strains may lead to emergence of novel BTV lineages, strains or serotypes with increased virulence of bluetongue outbreaks. The introduction of BTV western stains/ genome segments could potentially be linked to animal movements. India has imported Corriedale, Merino and Dorset sheep from Australia and Rambouillet sheep from America during 1970s and 1980s (Jain et al., 1986; Prasad et al., 2009; Maan et al., 2012f) and cattle from Belgium, France, Germany, Nepal, South Africa, Russia, the United Kingdom and the United States (Rao et al., 2012) during 2002 to 2005. Some of these countries used live attenuated vaccines during these periods. The vaccine virus can cause significant levels of viraemia and have possibility of transmission by vector or potential for reversion to virulent strain or reassortment with other field or vaccine strain. In addition to import of vaccinated animals, unauthorized movement of vaccine viruses, despite trade restriction, cannot be ruled out.
Seg–5 of VJW64 isolate showed only 88.5 – 88.6% sequence identity with western BTV10 vaccine strain and western BTV2 (IND2003/02) strain circulating in India, and is therefore not derived from these isolates. However high sequence identity (98.4%) of VJW64 isolate with US isolate prototype 600565 and reassortant strain (BTV2, BTV3 and BTV23) detected in India indicated its possible source from one of these isolates and wider circulation of this western genome segment–5.
Sequence analysis and phylogenetic relationship indicate that BTV strains in different geographical regions have evolved over long period of time by acquiring gene mutations, homologous recombination and genome segment reassortment. Further, movement of viral strains from one geographical region to other and thereby circulation provides opportunities for generation of new strains best suited to the climatic conditions. This led to development of regional variants of each genome segment. India being a tropical country has different geo–climatic conditions. In order to gain a selective advantage and to meet the new challenges of new ecosystem including vertebrate and non–vertebrate hosts, the virus tends to undergo gene reassortment event. The presence of western genome seg–5 in so many Indian BTV isolates (VJW64/08/IND, IND1982/01, IND2003/08, and IND1988/02) seems to provide an advantage to these isolate and explain their wider circulation and increased virulence during the outbreak.
Although BTV genome segment reassortment between eastern and western topotypes has previously been reported in BTV2 (Maan et al., 2012c), BTV3 (Maan et al., 2012a) and BTV23 (Maan et al., 2012e) serotypes from India, present study reports the occurrence of genetic reassortment in isolate of BTV16 serotype for the first time in India.
CONCLUSION
In the present study we sequenced full length genome seg–5 of Indian isolate (VJW64) of BTV16 serotype and carried out phylogenetic comparison with other global isolates of bluetongue virus. It was found that seg–5 of this isolate has nucleotide sequence identity of less than 80% from seg–5 of eastern topotypes and more than 88.5% from seg–5 of western topotypes. However, closer relationship of seg–2 and seg–6 of this isolate (Shafiq et al., 2013) with eastern topotypes reflect that virus has derived segments from both topotypes and is a reassortant. The possibility of intercontinental movement of viral isolates and segments are well documented along with earlier report of circulation of western isolates in India. Further a wide spread occurrence of this segment in various isolates and reassortants reflects its selective advantage. It was probably one of the reasons for the involvement of these isolates in severe outbreaks of bluetongue in India. The report describes the occurrence of a reassortant of BTV16 serotype for the first time in India.
ACKNOWLEDGEMENTS
The financial support for the present study was provided by Indian Council of Agricultural Research, New Delhi under ‘All India network programme on Bluetongue’ and Haryana State Council for Science and Technology, Panchkula. The study has taken some data from public database. We thank these sequence submitters. We are also thankful to Department of Animal Biotechnology, College of Veterinary Sciences, LLR University of Veterinary and Animal Sciences, Hisar for providing infrastructural facilities.
CONFLICT OF INTEREST
There is no conflict of interest regarding the present study.
REFERENCES
Balasuriya UB, Nadler SA, Wilson WC, Pritchard LI, Smythe AB, Savini G, Monaco F, Santis DP, Zhang N, Tabachnick WJ and Maclachlan NJ (2008). The NS3 proteins of global strains of bluetongue virus evolve into regional topotypes through negative (purifying) selection. Vet. Microbiol. 126: 91–100.
http://dx.doi.org/10.1016/j.vetmic.2007.07.006
PMid:17706379
Batten CA, Maan S, Shaw AE, Maan NS and Mertens PP (2008). A European field strain of bluetongue virus derived from two parental vaccine strains by genome segment reassortment. Virus Res. 137: 56–63.
http://dx.doi.org/10.1016/j.virusres.2008.05.016
PMid:18598726
Belhouchet M, Jaafar FM, Firth AE, Grimes JM, Mertens PPC and Attoui H (2011). Detection of a fourth orbivirus non–structural protein. PLoS One. 6: e25697.
http://dx.doi.org/10.1371/journal.pone.0025697
PMid:22022432 PMCid:PMC3192121
Carpi G, Holmes EC and Kitchen A (2010). The evolutionary dynamics of bluetongue virus. J Mol. Evol. 70: 583–592.
http://dx.doi.org/10.1007/s00239-010-9354-y
PMid:20526713
Cowley JA and Gorman BM (1989). Cross–neutralization of genetic reassortants of bluetongue virus serotype 20 and 21. Vet. Microbiol. 19: 37–51.
http://dx.doi.org/10.1016/0378-1135(89)90089-8
Gorman BM (1990). The bluetongue viruses. Curr. Top. Microbiol. Immunol. 162: 1–19.
http://dx.doi.org/10.1007/978-3-642-75247-6_1
Gould AR and Pritchard LI (1990). Relationships amongst bluetongue viruses revealed by comparisons of capsid and outer coat protein nucleotide sequences. Virus Res. 17: 31–52.
http://dx.doi.org/10.1016/0168-1702(90)90078-P
Hall TA (1999). BioEdit: A user friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symposium, Series 41, 95–98.
He CQ, Ding NZ, He M, Li SN, Wang XM, He HB, Liu XF and Guo HS (2010). Intragenic recombination as a mechanism of genetic diversity in bluetongue virus. J. Virol. 84: 11487–11495.
http://dx.doi.org/10.1128/JVI.00889-10
PMid:20702614 PMCid:PMC2953192
Hofmann MA, Renzullo S, Mader M, Chaignat V, Worwa G and Thuer B (2008). Genetic characterization of Toggenburg orbivirus, a new bluetongue virus, from goats, Switzerland. Emerg. Infect. Dis. 14: 1855–1861.
http://dx.doi.org/10.3201/eid1412.080818
PMid:19046507 PMCid:PMC2634640
Huismans H, Van Dijk AA and Els JH (1987). Uncoating of parental bluetongue virus to core and subcore particles in infected L cells. Virology. 157: 180–188.
http://dx.doi.org/10.1016/0042-6822(87)90327-8
Jain NC, Sharma R and Prasad G (1986). Isolation of bluetongue virus from sheep in India. Vet. Record. 119: 17–18.
http://dx.doi.org/10.1136/vr.119.1.17
PMid:3018987
Laemmli UK (1970). Cleavage of structural proteins during the assembly of head of bacteriophage T4. Nature. 227: 680–685.
http://dx.doi.org/10.1038/227680a0
PMid:5432063
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ and Higgins DG (2007). Clustal W and Clustal X version 2.0. Bioinformatics. 23(21): 2947–2948.
http://dx.doi.org/10.1093/bioinformatics/btm404
PMid:17846036
Maan NS, Maan S, Guimera M, Nomikou K, Morecraft E, Pullinger G, Belaganahalli MN and Mertens PPC (2012a). The genome sequence of a reassortant bluetongue virus serotype 3 from India. J. Virol. 86: 6375–6376.
http://dx.doi.org/10.1128/JVI.00536-12
http://dx.doi.org/10.1128/JVI.00420-12
http://dx.doi.org/10.1128/JVI.00671-12
http://dx.doi.org/10.1128/JVI.00731-12
http://dx.doi.org/10.1128/JVI.01672-12
http://dx.doi.org/10.1128/JVI.00188-12
http://dx.doi.org/10.1128/JVI.00596-12
Maan NS, Maan S, Guimera M, Pullinger G, Singh KP, Nomikou K, Belaganahalli MN and Mertens PP (2012b). Complete genome sequence of an isolate of bluetongue virus serotype 2, demonstrating circulation of a Western topotype in southern India. J. Virol. 86: 5404–5405.
http://dx.doi.org/10.1128/JVI.00536-12
http://dx.doi.org/10.1128/JVI.00420-12
http://dx.doi.org/10.1128/JVI.00731-12
http://dx.doi.org/10.1128/JVI.00671-12
http://dx.doi.org/10.1128/JVI.01672-12
http://dx.doi.org/10.1128/JVI.00188-12
http://dx.doi.org/10.1128/JVI.00596-12
Maan NS, Maan S, Nomikou K, Guimera M, Pullinger G, Singh KP, Belaganahalli MN and Mertens PPC (2012c). The genome sequence of bluetongue virus type 2 from India: evidence for reassortment between eastern and western topotype field strains. J. Virol. 86: 5967–5968.
http://dx.doi.org/10.1128/JVI.00536-12
http://dx.doi.org/10.1128/JVI.00671-12
http://dx.doi.org/10.1128/JVI.00731-12
http://dx.doi.org/10.1128/JVI.00188-12
http://dx.doi.org/10.1128/JVI.00596-12
Maan NS, Maan S, Nomikou K, Prasad G, Singh KP, Belaganahalli MN and Mertens PP (2012d). Full genome sequence of bluetongue virus serotype 1 from India. J. Virol. 86: 4717–4718.
http://dx.doi.org/10.1128/JVI.00536-12
http://dx.doi.org/10.1128/JVI.00420-12
http://dx.doi.org/10.1128/JVI.00671-12
http://dx.doi.org/10.1128/JVI.00731-12
http://dx.doi.org/10.1128/JVI.01672-12
http://dx.doi.org/10.1128/JVI.00188-12
http://dx.doi.org/10.1128/JVI.00596-12
Maan S, Maan NS, Guimera M, Nomikou K, Singh KP, Pullinger G, Belaganahalli MN and Mertens PP (2012e). Genome sequence of a reassortants strain of bluetongue virus serotype 23 from western India. J. Virol. 86: 7011–7012.
http://dx.doi.org/10.1128/JVI.00536-12
http://dx.doi.org/10.1128/JVI.00420-12
http://dx.doi.org/10.1128/JVI.00671-12
http://dx.doi.org/10.1128/JVI.00731-12
http://dx.doi.org/10.1128/JVI.01672-12
http://dx.doi.org/10.1128/JVI.00188-12
http://dx.doi.org/10.1128/JVI.00596-12
Maan S, Maan NS, Mertens PPC, Nomikou K and Belaganahalli MN (2012f). Reply to intercontinental movement of bluetongue virus and potential consequences to trade. J. Virol. 86: 8342–8343.
http://dx.doi.org/10.1128/JVI.01024-12
PMCid:PMC3421692
Maan S, Maan NS, Nomikou K, Batten C, Antony F, Belaganahalli MN, Samy AM, Reda AA, Rashid SA, Batel ME, Oura CAL and Mertens PPC (2011). Novel bluetongue virus serotype from Kuwait. Emerg. Infect. Dis. 17: 886– 889.
http://dx.doi.org/10.3201/eid1705.101742
PMid:21529403 PMCid:PMC3321788
Maan S, Maan NS, Pullinger G, Nomikou K, Morecraft E, Guimera M, Belaganahalli MN and Mertens PPC (2012g). The genome sequence of bluetongue virus type 10 from India: evidence for circulation of a western topotype vaccine strain. J. Virol. 86: 5971–5972.
http://dx.doi.org/10.1128/JVI.00536-12
http://dx.doi.org/10.1128/JVI.00420-12
http://dx.doi.org/10.1128/JVI.00671-12
http://dx.doi.org/10.1128/JVI.00731-12
http://dx.doi.org/10.1128/JVI.00188-12
http://dx.doi.org/10.1128/JVI.00596-12
PMid:22532535 PMCid:PMC3347277
Maan S, Maan NS, Smith RN, Batten CA, Shaw AE, Anthony SJ, Samuel AR, Darpel KE, Veronesi E, Oura CA, Singh KP, Nomikou K, Potgieter AC, Attoui H, Rooji VE, Rijn VP, Clercq DK, Vandenbussche F, Zientara S, Breard E, Sailleau C, Beer M, Hoffman B, Mellor PS and Mertens PP (2008). Sequence analysis of bluetongue virus serotype 8 from the Netherlands 2006 and comparison to other European strains. Virology. 377: 308–318.
http://dx.doi.org/10.1016/j.virol.2008.04.028
PMid:18570969
Mertens PP, Maan NS, Prasad G, Samuel AR, Shaw AE, Potgieter AC, Anthony SJ and Maan S (2007). Design of primers and use of RT–PCR assays for typing European bluetongue virus isolates: differentiation of field and vaccine strains. J. Gen. Virol. 88: 2811.
http://dx.doi.org/10.1099/vir.0.83023-0
PMid:17872535
Mertens PP, Pedley S, Cowley JA, Burrough JN, Corteyn AH, Jeggo MH, Jennings DM and Gorman BM (1989). Analysis of the roles of bluetongue virus outer capsid proteins VP2 and VP5 in determination of virus serotype. Virology. 170: 561–565.
http://dx.doi.org/10.1016/0042-6822(89)90447-9
Mertens PPC, Maan NS, Nomikou K, Samuel A, Shaw A and Maan S (2008). Molecular epidemiology studies of bluetongue virus. In: Mertens PPC, Baylis M, Mellor P (Eds.), Bluetongue Virus. Elsevier, New York, pp. 135–166.
PMid:18535825
Minakshi P, Singh R, Ranjan K, Kumar P, Joshi CG, Reddy YK and Prasad G (2012). Complete genome sequence of bluetongue virus serotype of goat origin from India. J. Virol. 86: 8337–8338.
http://dx.doi.org/10.1128/JVI.01128-12
PMid:22787269 PMCid:PMC3421637
Nomikou K, Dovas CI, Maan S, Anthony SJ, Samual AR, Papanastassopoulou M, Maan NS, Mangana O and Mertens PP (2009). Evolution and phylogenetic analysis of full–length VP3 genes of Eastern Mediterranean bluetongue virus isolates. PLoS One. 4: e6437.
http://dx.doi.org/10.1371/journal.pone.0006437
PMid:19649272 PMCid:PMC2713410
Prasad G (2000). Wither bluetongue research in India. Indian J. Microbiol. 40: 163–175.
Prasad G, Sreenivasulu D, Singh KP, Mertens PPC and Maan S (2009). Bluetongue in the Indian subcontinent, p 167–196. In Mellor PS, Baylis M, Mertens PPC (eds.), Bluetongue, 1st ed. Elsevier/ Academic Press, London, United Kingdom.
http://dx.doi.org/10.1016/B978-012369368-6.50012-5
PMid:19339108
Rao PP, Hedge NR and Reddy YN (2012). Intercontinental movement of bluetongue virus and potential consequences to trade. J. Virol. 86: 8341.
http://dx.doi.org/10.1128/JVI.00968-12
PMid:22787271 PMCid:PMC3421687
Sambrook J, Russell DW and Jassen AK (2001). Molecular cloning: A laboratory manual. Cold Spring Harbour Laboratory Press, New York.
Savini G, Hamers C, Conte A, Mingliaccio P, Bonfini B, Teodori L, Ventura DM, Hudelet P, Schumacher C and Caporale V (2009). Assessment of efficacy of a bivalent BTV–2 and BTV–4 inactivated vaccine by vaccination and challenge in cattle. Vet. Microbiol. 133: 1–8.
http://dx.doi.org/10.1016/j.vetmic.2008.05.032
PMid:18617335
Shafiq M, Minakshi P, Bhateja A, Ranjan K and Prasad G (2013). Evidence of genetic reassortment between Indian isolate of bluetongue virus serotype 21 (BTV–21) and bluetongue virus serotype 16 (BTV–16). Virus Res. 173: 336–343.
http://dx.doi.org/10.1016/j.virusres.2013.01.009
PMid:23353779
Shaw AE, Ratinier M, Nunes SF, Nomikou K, Caporale M, Golder M, Allan K, Hamers C, Hudelet P, Zientara S, Breard E, Mertens P and Palmarini M (2013). Reassortment between two serologically unrelated bluetongue virus strains is flexible and can involve any genome segment. J. Virol. 87: 543–557.
http://dx.doi.org/10.1128/JVI.02266-12
PMid:23097432 PMCid:PMC3536370
Singh KP, Maan S, Samuel AR, Rao S, Meyer A and Mertens PPC (2004). Phylogenetic analysis of bluetongue virus genome segment 6 (encoding VP5) from different serotypes. Vet. Ital. 40: 479–483.
PMid:20422573
Svensson L, Uhnoo I, Grandien M and Wadeli G (1986). Molecular epidemiology of rotavirus infections in Upsala, Sweden, 1981; disappearance of a predominant electropherotype. J. Med. Virol. 18: 101–111.
http://dx.doi.org/10.1002/jmv.1890180202
PMid:3005484
Tamura K, Peterson D, Peterson N, Stecher G, Nei M and Kumar S (2011). MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance and maximum parsimony methods. Mol. Biol. Evol. 28(10): 2731–2739.
http://dx.doi.org/10.1093/molbev/msr121
PMid:21546353 PMCid:PMC3203626
Veronesi E, Hamblin C and Mellor PS (2005). Live attenuated bluetongue vaccine viruses in Dorset Poll sheep, before and after passage in vector midges (Diptera: Ceratopogonidae). Vaccine. 23: 5509–5516.
http://dx.doi.org/10.1016/j.vaccine.2005.07.039
PMid:16111787

