
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 2 (7): 401 – 406Biology of Embryo Development in Pigeon Columba livia domesticus in Conditions of Artificial Incubation
Monika Łukasiewicz1*, Karolina Wnęk2, Karolina Boruc1
- Faculty of Animal Sciences, Department of Poultry Breeding, Warsaw University of Life Sciences (SGGW), Warsaw, Poland
- Faculty of Animal Sciences, Cattle Breeding Division, Warsaw University of Life Sciences (SGGW), Warsaw, Poland
*Corresponding author:monika_lukasiewicz@sggw.pl
ARTICLE CITATION:
Łukasiewicz M, Wnęk K, Boruc K (2014). Biology of embryo development in pigeon Columba livia domesticus in conditions of artificial incubation. Adv. Anim. Vet. Sci. 2 (7): 401 – 406.
Received: 2014–06–24, Revised: 2014–08–14, Accepted: 2014–08–15
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/journal.aavs/2014/2.7.401.406
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
The study was conducted on the eggs of pigeon Columba livia domesticus collected in an amateur pigeon breeding facility during the breeding season. Eggs were collected in the morning from the long–beaked pigeons to make sure that they have been laid recently. They were incubated at 37.8oC and humidity of 65% (standard conditions for the development of the domestic chicken embryos). A total of nearly 70 embryos have been analyzed at different stages of development. 4 fertilized eggs were opened every day and analyzed for external morphological features according to the Hamburger and Hamilton key (1951). It was found that the embryonic development of pigeon Columba livia domesticus was shorter (17 days) by about 4 days compared with embryos of domestic chicken Gallus gallus domesticus (21 days). Further, it was observed that the pigeon embryos developed slower in the first 14 days of embryogenesis, and a marked acceleration of their development occurred from day 14 of incubation. Artificial incubation enables successful raising of Columba livia domesticus pigeon chicks. The remaining period of the growth and development of chicks is possible by placing them with foster parents, long–beaked pigeons.
INTRODUCTION
The rock dove (Columba livia), living in southern Europe, southern and central Asia, and northern Africa is considered an ancestor of domestic pigeons. The gray–blue coloring of this pigeon is similar to that of homing pigeon (Milan and Slavibor, 1995). The wild rock dove is medium in size and resembles the so–called feral urban pigeon, present almost all over the globe. Domestic pigeon is a species, which domestication was a success at the turn of the century. Pigeon breeding belongs to the ancient human activities, and over time it acquired a completely different character, i.e. ornamental and sporting use of these birds. At the beginning they were kept on religious grounds, and only later used for postal purposes (Nowicki et al., 1996). The harvest and storage of cereal crops caused the wild animals to search for the human households where they could easily find food. People also benefited from the presence of these birds, e.g., in the form of eggs. Man was accompanied by a pigeon throughout the development of civilization. When people began to build the cities with tall buildings and towers that resembled natural nesting places of these species such as rocks, the birds began to settle there and the population increased (Tomiałojć and Stawarczyk, 2003). In some cultures, pigeons were considered sacred, because it was claimed that God itself sent them down on church towers. Pigeon has played a very important role in Christian symbolism. The symbol of this bird is often encountered in the catacombs. It is in the form of a dove that people imagined the soul after death reaching into heaven, similarly as the figure of the Holy Spirit descending upon the earth (Herman, 1963). Taylor (2006) wrote that in the Old Testament, pigeons were one of the main birds of sacrifice, but in the New Testament they were primarily associated with the baptism of Christ. Pigeons are also a symbol of hope, new life, peace and abundance, spiritual and material goods, and the forgiveness that God sent down to the Noah's Ark under the symbol of a bird (Cooper, 1998). The Islamic religion considers this bird to be holy, because it protected Muhammad during his escape (Herder, 1992). Litwiński (1995) reported that some of the earliest information on the breeding of these birds came from the third and fourth centuries BC from the land of Egypt. Reliefs have been discovered in the tombs of the pharaohs, which depicted scenes with pigeons, e.g., the release of homing pigeons or doves receiving nourishment. Cypriot excavations of dovecotes created on the model of small chapels are dated to the same time period (Herman, 1963). This species and its breeding spread to areas of current Italy, Spain and Greece with the expansion of the Phoenicians in the Mediterranean, where to this day paintings can be find with the image of a dove. Peterfi (1977) reports that thanks to the Greeks domestic pigeon spread across the area of Italy of that times, where it was called "colymbos", hence the Latin name columba of this bird. In the first century BC the Roman scholar Varro has described for the first time the then breeding of pigeons, in his famous work "Rerum rusticarum". He listed the rock dove, common wood pigeon, and described the dovecotes, rearing of young birds, feeding and their local market price. Pigeons have been used to convey messages due to the incredible "sense of navigation" and the speed and fluidity of the flight. Dovecotes were also installed on fishing boats, where the birds were sent with information about the shoals of fish. Numerous military expeditions and commercial communication widened the worship and range of pigeon breeding facilities, also in Asia, where the famous scholar Avicenna, in his work "Canon Medicinae'' of the eleventh century, mentioned the dietary value of meat of these birds. This species came to Europe via Arab nations.
The aim of the study was to analyze the biology (preparation of the key) of embryo development of pigeons Columba livia domesticus in conditions of artificial incubation.
MATERIALS AND METHODS
Description of Experimental Material
Eggs were collected in an amateur pigeon breeding facility during the breeding season. There were two groups of birds, i.e., long–beaked and short–beaked pigeons (without the possibility of feeding their offspring). For this study, eggs were collected in the morning from long–beaked pigeons, serving as foster parents for short–beaked pigeons to make sure that they have been recently laid, planting them at the same time the eggs of short–beaked pigeons.
Incubation
The eggs were incubated at 37.8oC and humidity of 65% (standard conditions for the development of the domestic chicken embryos) in a hatching incubator (Heka, Germany) equipped with a module controlling the temperature, humidity and egg–turning system. The apparatus had insulated glazing, interior lighting, automatic egg turning, 3 breeding drawers for selected species of birds, 2 hatching drawers and 5 liter canister of water capable of maintaining the preset humidity.
Prior to placing the eggs into the incubator, they were weighed individually on a technical scale to the nearest 0.01 g and then x–rayed and re–weighed weekly to determine the weight loss of eggs.
Embryo Analysis
A total of nearly 70 embryos have been analyzed at different stages of development. 4 fertilized eggs were opened every day and analyzed for external morphological features according to the Hamburger and Hamilton key (1951). The eggs were taken out of the incubator after 12 h, (a single batch of eggs had 4–6 pieces) and left at about 17–18oC in order to cool the surface of the shells. This was aimed at slowing down the development of embryos, which were positioned just underneath the eggshell membranes and preventing possible differences in their morphological assessment.
Eggs were opened using the method described by Callebaut (1981), involving the creation of an artificial air chamber over the embryo. Blastoderm was stained by the introduction of India ink (KOH–I–NOOR Hardtmuth) into the space under the embryo. The surface of the blastodisc was then fixed with 1% acetic acid, and the preparation was observed under binocular at 25x magnification.
Photographic documentation was made with the use of microscope Nikon Model Eclipse E 200 MV RS Tokyo, Japan with built–in camera. Images were taken at 60x magnification and in some cases at 10x and 200x. Additionally, a digital camera Canon EOS 50D was used. Image processing was performed using Microsoft Photo Editor.
The material analyzed has become part of the didactic material of embryology at the Department of Poultry Breeding.
RESULTS AND DISCUSSION
Biological Analysis of the Clutch
All bird species are egg–laying animals. The size and shape of the egg as well as the color of the shell and the number of eggs in the clutch is specific and often quite different for various species of birds (Christians, 2002). As it might be expected, the size of the egg is approximately related to the size of the bird. The size of the clutch is influenced by factors such as natural selection, or the ability to feed a certain number of young, which particular species can successfully raise without fear of malnutrition due to the necessity of division of existing food supplies among a large number of hungry chicks. The successive phases of the reproductive cycle in pigeons comes so rapidly that often a female laying eggs to the next nest can be observed, while the male is still feeding young from the first clutch. The variability of the number of hatches in different species and the associated number of young raised in one breeding season, are a reflection of fundamental differences in growth and survival chances of the young birds.
Average weight of eggs of pigeons in the experiment was 15.71 g. of the farm birds only quail eggs have a lower weight (11.3 g) (Kul and Seker, 2004; Ayasan et al., 2006; Adamski, 2008). The tiny hummingbird lays the smallest eggs, weighing less than 1 g, while the ostrich lays the largest. The size of an egg is also to some extent dependent on the amount of nutrients stored therein, and the type and degree of care that developing chicks receive from their parents.
The egg weight control was carried out in this study in order to determine their weight loss. During the incubation, the eggs were losing their initial weight, mainly due to the loss of water. The average egg weight difference between the beginning and the end of the incubation was 3.02 g. Egg weight loss was very diverse, which resulted from large differences in the initial weight of the eggs tested (Table 1). It was found that the average weight loss of incubated eggs of pigeons for the whole incubation period was 18.14%, which is much higher compared to the domestic chicken eggs. According to Borzemska (1996), during normal hatches, hen's egg is losing about 12% of its mass after 18 days of incubation.
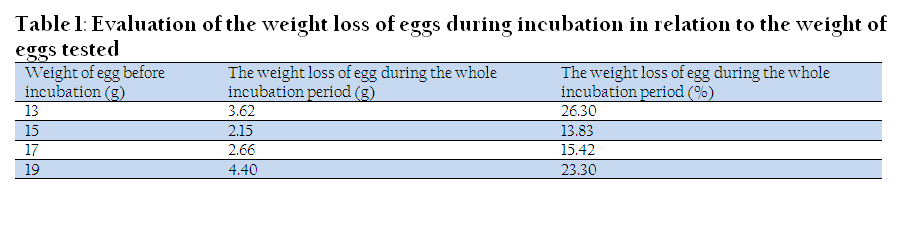
Table 1: Evaluation of the weight loss of eggs during incubation in relation to the weight of eggs tested
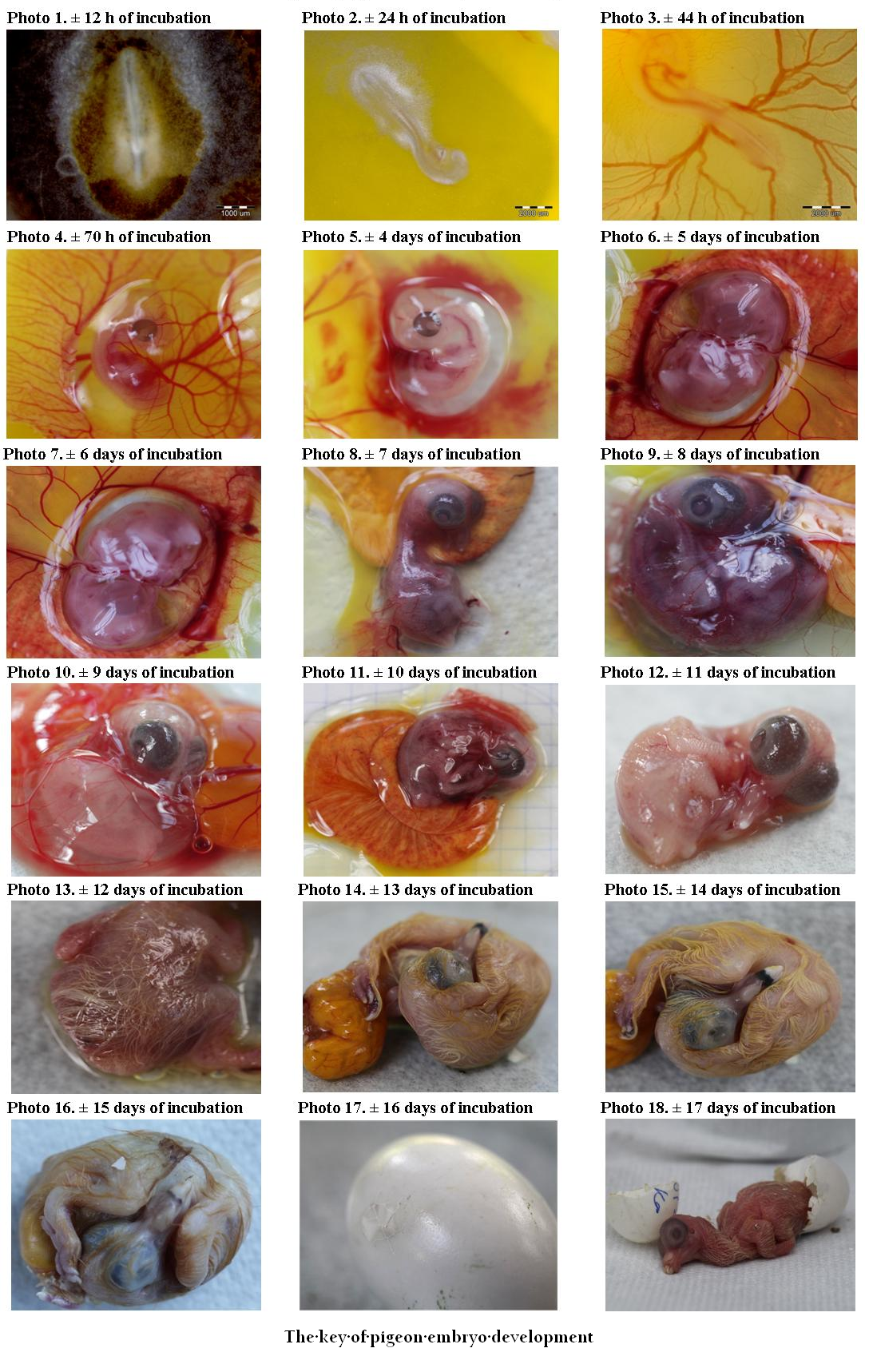
The Key of Pigeon Embryo Development
Photo 1 (± 12 h of incubation): Formation of the primitive streak, which gives rise to the notochord
Photo 2 (± 24 h of incubation): 5 pairs of somites. Brain vesicles, the rudiments of the head, are clearly differentiated. Fetal membranes are formed. Network of blood vessels is visible on the front area of the yolk membrane. Rudiments of the limbs begin to form as well as eye vesicles, ear cavities, heart and arteries.
Photo 3 (± 44 h of incubation): 25 pairs of somites. Brain vesicles start to differentiate, midbrain is much more prominent than the other vesicles of the brain. Clear eye pigmentation can be seen. The embryo assumes the shape of a letter C and has a large head. Midbrain is well developed. Limbs continue to form and the skeleton starts to form, lungs start to develop. The yolk membrane is covered to about half the mass of the yolk with a well–developed network of arteries and veins. First heartbeats appear.
Photo 4 (± 70 h of incubation): 40–43 pairs of somites. The beak starts to form, the maxillary part of the beak is longer than the mandible portion. The rudiments of wings. Segmentation of the limbs begins. The heart with visible chambers. Allantois larger and well vascularized.
Photo 5 (4 days of incubation): The embryo becomes white, inner structures less visible. Large eyes. A clear development of the beak. Allantois completely encloses the embryo. Limb buds more developed.
Photo 6 (5 days of incubation): Allantois larger than in the previous stage. Further development of the beak. Longer limbs with palpable nodules – scales. Formation of reproductive organs and sexual differentiation. Vasculature entirely covers the yolk mass.
Photo 7 (6 days of incubation): Clearly visible large eyes. Extension of the limbs, the fore and hind limbs have indentations. The thigh, lower leg and foot develop, and in wing: the arm and forearm.
Photo 8 (7 days of incubation): The upper part of the beak is longer and bent downwards. Interdigital spaces appear in the wings. Toes begin to form. Elongation of the neck. Rudiments of primary feathers appear.
Photo 9 (8 days of incubation): Clear hardening of the beak. Longer wings. Hind limbs transparent and longer than the wings. Limbs noticeably more developed than in the previous stage. The eyelids are visible around the eyeball.
Photo 10 (9 days of incubation): Further elongation of the beak. Ear cavities are visible. Further development of the eyeball. Eyelids cover half of the eyeballs. Limbs are developed. Feather follicles visible on almost the entire body with the exception of abdomen and neck.
Photo 11 (10 days of incubation): Hind leg digits clearly separated. Enlargement of feather follicles. They cover the entire body with the exception of the abdomen. The upper and lower eyelids cover the sclera. The embryo is immersed in the yolk, allantois contacts the shell through the chorion. Filamentous feathers grow from the feather follicles in the wings and femoral part. Claws appear.
Photo 12 (11 days of incubation): Beak proportionally elongated with the visible white protuberance called the egg tooth. Tips of the claws become white. Feathers similar to the previous stage.
Photo 13 (12 days of incubation): The long filamentous feathers in the trunk area, short on the head, neck and wings. Visible pattern of scales on tarsi. Eyelids cover most of the eye.
Photo 14 (13 days of incubation): Hardening of the beak. Filamentous feathers longer than in the previous stage. Tarsi covered with granular scales. The eyelids almost completely cover the eyeballs. Claws become white.
Photo 15 (14 days of incubation): Eyelids closed but not fused. All the other developmental stages similar to the previous ones. Elongation of filamentous down feathers. Outgrowth over the nostrils is clearly marked.
Photo 16 (15 days of incubation): Beak rotates towards the air cell. The eyelids are the same as in the previous stage.
Photo 17 (16 days of incubation): Yolk sac is pulled into the abdominal cavity of the embryo. Filamentous feathers become yellow. The embryo occupies virtually the whole space of the egg. Puncturing the shell around the air cell, the allantois is in atrophy.
Photo 18 (± 17 days): Embryos have closed eyelids, and begin to hatch.
There have been many observations carried out on the course of hatching and quality of shells in various species of wild and domestic birds (Ar et al., 1979; Burton and Tullett, 1983; Tullett, 1984; Roberts et al., 1995; Mróz, 1998; Lourens at al., 2005). The fundamental factors affecting the quality of breeding in wild birds include the temperature, location of the nest, its type and the occurrence of environmental pollution as well as the fitness of the birds participating in breeding (Skutch, 1962; Olsen, 1982; Kwieciński et al., 2009; Olea et al., 2012). Number of eggs laid by a female is also dependent on the species, age and size of the bird, and can range from one to a dozen eggs. The length of the artificial or natural incubation, to a large extent is associated with the rate of development of the embryo in the egg. This process is determined by the active temperature control of eggs heated, which is designed to protect them from overheating or overcooling (Grant, 1982; Haftron, 1984; Webb, 1987; Conway and Martin, 2000; Albokhadaim et al., 2010; Ekine and Ajuogu, 2011; Bergoug et al., 2013; Maatjens et al., 2014a; Maatjens et al., 2014b). However, the basic and common feature of all eggs is their structure and function they perform for the developing avian embryo. Birds belong to amniotes, which embryos are enclosed by fetal membranes: allantois, collecting unnecessary and harmful products of metabolism, amnion, enclosing the embryo, thereby creating an intrauterine–like life environment, and chorion, participating in the processes such as gas exchange (Duncker, 1978; Hoyt, 1979). During the first day of incubation, a line can be noticed called the primitive streak, which allows the formation of a third layer of cells, from which organs of the embryo are formed. These cells form three layers: ectoderm, mesoderm and endoderm. The following structures are formed from the first layer: feathers, skin, beak, nervous system, claws, oral cavity and eyes. From the mesoderm develops structure such as skeleton and muscles, and from endoderm respiratory organs, and organs of the digestive and secretion systems. At the end of the first days of incubation the head, eyes, nervous system and neural tube begin to form. Heart, depending on the species, develops on the second day, and starts to function after about 44 hours of incubation (Noden, 1988; Garcia–Martinez and Schoenwolf, 1993; Wei and Mikawa, 2000; Inagaki et al., 1993).
During incubation, the eggs lose their initial weight due to water loss progressing with the development of the embryo. The weight loss of eggs is usually quite different and varies between species. It can inform about the proper development of the embryo and/or about its cessation. The initial mass of eggs affects the hatching rate (Majewska et al., 2010).
In the final days of embryonic development, the chick starts to try breathing with lungs, which contributes to the contraction of the neck muscles and progressive shell puncturing, and ultimate getting out of it. Chick has the egg tooth, which significantly simplifies the process of hatching. Honza et al., (2001) demonstrated in their study that the method of puncturing the egg with the egg tooth as well as the way of escape of chicks from eggs vary for the different species of birds. Although significant differences exists in the clutches, resulting, among others, from specific environments, nesting, or the size and weight of the birds, affecting reproductive success, an egg plays the basic and essential function in reproduction that allows the normal development of the avian embryo.
CONCLUSIONS
The results allowed the formulation of the conclusions, which may be important for amateur pigeon breeding. The diverse and often attractive appearance of these birds causes the interest of breeders and breeding is a key to the success in reproduction.
Although the embryonic development of the pigeon Columba livia domesticus is shorter (17 days) by approximately 4 days compared with embryos of domestic chicken Gallus gallus domesticus (21 days), it was observed that the embryos developed more slowly in pigeons in the first 14 days of embryogenesis. However, a marked acceleration of their development occurred from day 14 of incubation.
Artificial incubation enables successful raising of chicks of Columba livia domesticus pigeon. Further process of the growth and development of chicks hatched is possible by placing them with foster parents, long–beaked pigeons.
Pigeon chicks obtained by artificial incubation will influence basic measures aimed at protecting the biodiversity, which under the Convention means "the diversity of all living organisms on Earth in terrestrial, marine and freshwater ecosystems as well as the ecological complexes, of which they are part; this includes diversity within species, between species and diversity of ecosystems", and will affect the protection of the Polish pigeon breeds threatened with extinction.
REFERENCES
Adamski M (2008). Zeszyty naukowe nr (rozprawy) zależności między składem morfologicznym jaj a wylęgowością piskląt wybranych gatunków ptaków. Wydawnictwo Uczelniane Uniwersytetu Technologiczno–Przyrodniczego, Bydgoszcz [in Polish].
Albokhadaim IF, Ismail IB, El–Bahr SM, Althnaian T (2010). The Effect of Manipulations of Incubation Temperature on Embryonic and Post–hatching Growth of Native Saudi Chickens. Pak. J Biol. Sci. 13: 852 – 856.
http://dx.doi.org/10.3923/pjbs.2010.852.856
PMid:21313911
Ar A, Rahn H, Paganelli CV (1979). The avian egg: mass and strength. Condor 81: 331 – 337.
http://dx.doi.org/10.2307/1366955
Ayasan T, Ozcan BD, Baylan M, Canogullari S (2006). The effects of dietary inclusion of probiotic protexin on egg yield parameters of japanese quails (Coturnix coturnix Japonica). Int. J. Poult. Sci. 5(8): 776 – 779.
http://dx.doi.org/10.3923/ijps.2006.776.779
Bergoug H, Burel C, Guinebretière M, Tong Q, Roulston N, Romanini CEB, Exadaktylos V, Mcgonnell IM, Demmers TGM, Verhelst R, Bahr C, Berckmans D, Eterradossi N (2013). Effect of pre–incubation and incubation conditions on hatchability, hatch time and hatch window, and effect of post–hatch handling on chick quality at placement. World's Poult. Sci. J. 69(2): 313 – 334.
http://dx.doi.org/10.1017/S0043933913000329
Borzemska WB (1996). Embriopatologia Drobiu. Drobiarstwo. 10: 5 – 9. [in Polish].
Burton FG, Tullett SG (1983). A comparison of the effects eggshell porosity on the respiration and growth of domestic fowl, duck and turkey embryos. Compar. Biochem. Physiol. 75 (2): 167 – 174.
http://dx.doi.org/10.1016/0300-9629(83)90065-8
Callebaut A (1981). A new method for making an artificial air space on top of fertilized eggs. Poultry Science, 60: 723 – 725.
http://dx.doi.org/10.3382/ps.0600723
PMid:7197784
Christians JK (2002). Avian egg size: variation within species and inflexibility within individuals. Biol. Rev. 77 (1): 1 – 26.
http://dx.doi.org/10.1017/S1464793101005784
PMid:11911371
Conway CJ, Martin TE (2000). Effects of ambient temperature on avian incubation behavior. Behav. Ecol. 11: 178 – 188.
http://dx.doi.org/10.1093/beheco/11.2.178
Cooper JC (1998). Zwierzęta Symboliczne i Mityczne, Dom Wydawniczy Rebis, Poznań [in Polish].
Duncker HR (1978). Development of the Avian Respiratory and Circulatory Systems. In: Piiper J (ed), Respiratory Function in Birds, Adult and Embryonic. Proc Life Sci. Springer–Verlag, Berlin Heidelberg New York. 260 – 273.
http://dx.doi.org/10.1007/978-3-642-66894-4_36
Ekine OA, Ajuogu PK (2011). The Impact of Pre–incubation Storage Temperatures on Hatchable Chicken Eggs. Innovations in Science and Engineering. 92 – 96.
Garcia–Martinez V, Schoenwolf GC (1993). Primitive–streak origin of the cardiovascular system in avian embryos. Dev. Biol. 159(2): 706 – 719.
http://dx.doi.org/10.1006/dbio.1993.1276
PMid:8405690
Grant GS (1982). Avian incubation. Egg temperature, nest humidity and behavioral thermoregulation in hot environment. Orn. Monogr. 30: 1 – 75.
Haftron S (1984). The behavior of an incubating female Coal Tit Parus ater in relation to experimental regulation of nest temperature. Fauna norv. Ser. 7: 12 – 20.
Hamburger V, Hamilton L (1951). A series of normal stages in the development of the chick embryo. J Morphol. 88: 49 – 92.
http://dx.doi.org/10.1002/jmor.1050880104
PMid:24539719
Herder F (1992). Leksykon Symboli, Tchu Dom Wydawniczy, Warszawa [in Polish].
Herman W (1963). Gołębie – ich rasy i hodowla. Państwowe Wydawnictwo Naukowe, Warszawa [in Polish].
Honza M, Picman J, Grim T, Novak V, Capek M, Mrlik W (2001). How to hatch from an egg of great structural strength. A study of the Common Cuckoo. J. Avian Biol. 32(3): 249 – 255.
http://dx.doi.org/10.1111/j.0908-8857.2001.320307.x
Hoyt DF (1979). Osmoregulation by avian embryos: the allantois functions like a toad's bladder. Physiol. Zool. 52(3): 354 – 362.
Inagaki T, Garcia–Martinez V, Schoenwolf GC (1993). Regulative ability of the prospective cardiogenic and vasculogenic areas of the primitive streak during avian gastrulation. Developmental Dynamics. 197(1): 57 – 68.
http://dx.doi.org/10.1002/aja.1001970106
PMid:8400411
Kul S, Seker I (2004). Phenotypic correlations between some external and internal egg quality traits in the japanese quail (Coturnix coturnix japonica). Int. J. Poult. Sci. 3(6): 400 – 405.
http://dx.doi.org/10.3923/ijps.2004.400.405
Kwieciński Z, Krawczyk A, Ćwiertnia P (2009). Wpływ temperatury otoczenia na wybrane elementy behawioru inkubacyjnego orła przedniego Aquila Chrysaetos w warunkach wolierowych. Notatki Ornitologiczne. 50: 1 – 8 [in Polish].
Litwiński R (1995). Gołębie rasowe. Państwowe Wydawnictwo Rolnicze i Leśne, Warszawa [in Polish].
Lourens A, van den Brand H, Meijerhof R, Kemp B (2005). Effect of eggshell temperature during incubation on embryo development, hatchability, and posthatch development. Poult Sci. 84(6): 914 – 20.
http://dx.doi.org/10.1093/ps/84.6.914
PMid:15971530
Maatjens CM, Reijrink IAM, Molenaar R, van der Pol CW, Kemp B, van den Brand H (2014a). Temperature and CO2 during the hatching phase. I. Effects on chick quality and organ development. Poultry Science. 93(3): 645 – 654. doi: 10.3382/ps.2013–03490.
http://dx.doi.org/10.3382/ps.2013-03490
Maatjens CM, Reijrink IAM, van den Anker I, Molenaar R, van der Pol CW, Kemp B, van den Brand H (2014b). Temperature and CO2 during the hatching phase. II. Effects on chicken embryo physiology. Poultry Science. 93(3): 655 – 663. doi: 10.3382/ps.2013–03491.
http://dx.doi.org/10.3382/ps.2013-03491
Majewska D, Ligocki M, Wołosiak K (2010). Masa jaja a wskaźnik wylęgowości strusi afrykańskich. Acta Sci. Pol. Zoot. 9 (4): 147 – 152 [in Polish].
Milan T, Slavibor P (1995). Leksykon Przyrody. Delta, Warszawa [in Polish].
PMCid:PMC2708290
Mróz E (1998). Studia nad zmiennością cech powierzchni skorupy jaj indyczych i ich związkiem z wylęgowością. Rozpr. habilitacyjna, Wyd. ART, Olsztyn [in Polish].
Noden DM (1988). Interactions and fates of avian craniofacial mesenchyme. Development. 103: 121 – 140.
PMid:3074905
Nowicki B, Pawlina E, Dubiel A (1996). Gołębie – chów, hodowla, rasy. Państwowe Wydawnictwo Rolnicze i Leśne, Warszawa, 9 – 70 [in Polish].
Olea G, Cienc Biol L, Sandoval M, Cienc L (2012). Embryonic development of Columba livia (Aves: Columbiformes) from an altricial–precocial perspective. Rev Colomb Cienc Pecu. 25: 3 – 13.
Olsen P (1982). Ecogeographic and temporal variation in the eggs and nests of the peregrine, falco peregrinus, (aves: falconidae) in Australia. Australian Wildlife Research. 9(2): 277 – 291.
http://dx.doi.org/10.1071/WR9820277
Peterfi S (1977). Hodowla gołębi. Państwowe Wydawnictwo Naukowe, Warszawa [in Polish].
Roberts JR, Brackpool CE, Solomon SE (1995). The ultrastructure of good and poor quality eggshells from australian layer strains. Proc. of the VI European Symposium on the Quality of Egg and Egg Products. Red. Ricardo Cepero Briz, 25–29 September 1995, Zaragoza, Spain. 107 – 115.
Skutch AF (1962). The constancy of incubation. Wilson Bulletin. 74: 115 – 152.
Taylor R (2006). Przewodnik po symbolice kościoła. Klub dla Ciebie, Warszawa [in Polish].
Tomiałojć L, Stawarczyk T (2003). Awifauna Polski. Rozmieszczenie, liczebność i zmiany, Warszawa [in Polish].
Tullett SG (1984). The porosity of avian eggshells. Comparative Biochem. Physiol. 78(1): 5 – 13.
http://dx.doi.org/10.1016/0300-9629(84)90083-5
Webb DR (1987). Thermal Tolerance of Avian Embryos: A Review. Condor. 89: 874 – 898.
http://dx.doi.org/10.2307/1368537
Wei Y, Mikawa T (2000). Formation of the avian primitive streak from spatially restricted blastoderm: evidence for polarized cell division in the elongating streak. Development. 127: 87 – 96.
PMid:10654603

