
South Asian Journal of Life Sciences


Review Article
South Asian Journal of Life Sciences. 1 (1): 5 – 11Angiosperm–Specific PcG Protein EMF1 Mainly Functions in Vegetative Development
Dong–Hong Chen1,2*, Yong Huang1,2, Chunlin Liu1, Ying Ruan1,2*
- Hunan Provincial Key Laboratory of Crop Germplasm Innovation and Utilization, Hunan Agricultural University, Changsha 410128, China
- Colleges of Bioscience and Biotechnology, Hunan Agricultural University, Changsha 410128, China
*Corresponding author:[email protected]; [email protected]
ARTICLE CITATION:
Chen D–H, Huang Y, Liu C and Ruan Y (2013). Angiosperm–specific PcG protein EMF1 mainly functions in vegetative development. South Asian J. Life Sci. 1 (1): 5 – 11.
Received: 2013–09–08, Revised: 2013–10–06, Accepted: 2013–10–07
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/25/108/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Polycomb group (PcG) proteins are relatively conserved in animal and plant, associating with the epigenetic modification and transcriptional silencing of target genes. Imaginably, PcG proteins in animal and plant should display different properties in some aspects as well, such as protein sequence, function, expression, and composition, based on both almost opposite behaviors. Here, we review a plant–specific PcG protein Embryonic Flower 1 (EMF1) which is necessary for maintenance of vegetative development. Sequence blast and phylogenic analysis indicated that EMF1 homologs are only found in angiosperm, divided into dicot and monocot groups. Domain searching showed that no obviously established domains are discovered in EMF1–like proteins. Then we introduce its expression pattern, subcellular localization and detailed functions both in diverse developmental stages and as a PcG role based on published results.
INTRODUCTION
Polycomb group (PcG) proteins were initially identified in Drosophila melanogaster, to repress the expression of the homeotic Hox genes related to embryo segmentation (Lewis, 1978; Jurgens, 1985). PcG proteins mainly form polycomb repressive complex 1 (PRC1) and PRC2, together acting as an epigenetic memory system that is conserved in plants and animals. In animal, the initial complex PRC2 catalyzes and introduces H3K27me3 into the target genes, and then the maintenance complex PRC1 recognizes and binds this epigenetic mark, and subsequently catalyzes histone H2A lysine119 monoubiquitylation (H2AK119ub) (Lund and van Lohuizen, 2004; Cernilogar and Orlando, 2005).
Drosophila core PRC2 components are comprised of Enhancer of Zeste [E(z), histone methyltransferase], Suppressor of Zeste12 [Su(z)12], and Extra Sex Combs (ESC), and p55. Correspondingly, Arabidopsis PRC2–like proteins include three E(z) homologs CLF, MEAEA (MEA), and SWINGER (SWN); three Su(z)12 homologs EMBRYONIC FLOWER2 (EMF2), FERTILIZATION–INDEPENDENT SEED2 (FIS2), and VERNALIZATION2 (VRN2); one Esc homolog FERTILIZATION–INDEPENDENT ENDOSPERM (FIE); five p55 homologs MULTICOPY SUPPRESSOR OF IRA1(MSI1)~5 (see review in Pien and Grossniklaus, 2007). These proteins can form different PRC2 complexes, which function at different development stages and events, but also share some target genes. The EMF2 complex (CLF/SWN, EMF2, FIE and MSI1) suppresses precocious flowering and cell dedifferentiation; the VRN2 complex (CLF/SWN, VRN2, FIE and MSI1) is responsible for FLC silencing after vernalization; and the FIS2 complex (MEA, SWN, FIS2, FIE and MSI1) prevents endosperm development in the absence of fertilization (see review by Hennig and Derkacheva., 2009).
Human core PRC1 components consist of BMI1, HPH1/EDR1, HPC2/CBX4, and RING1s (RING1A/RING1 and RING1B/RING2/RNF2), homologs of Drosophila Posterior sex combs (Psc), Polyhomeotic (Ph), Polycomb (Pc) and dRing1/Sce, respectively (Shao et al., 1999; Francis et al., 2001). Human BMI1, RING1A and RING1B all belong to Ring finger proteins, together forming an E3 ubiquitin ligase complex that monoubiquitinates histone H2A at lysine 119 (H2A–K119ub), where RING1B performs most of catalytic activity, RINGA only plays a minor role, but BMI1 alone does not show any E3–ligase activity and only enhances RING1B’s activity (Wang et al., 2004; Buchwald et al., 2006). PRC1 complex is highly conserved in plant as well (Molitor and Shen, 2013). In Arabidopsis, there are a Pc–functional equivalent, LIKE HETEROCHROMATIN PROTEIN1/TERMINAL FLOWER2 (LHP1/TFL2), two RING1 homologs, AtRING1a and AtRING1b; and three BMI1 homologs, AtBMI1a, AtBMI1b, and AtBMI1c (Zhang et al., 2007; Xu and Shen, 2008; Chen et al., 2010). It has been confirmed that AtRING1 proteins can interact with AtBMI1 proteins and LHP1 (Chen et al., 2010). Sequence homology showing Arabidopsis LHP1 encodes a HETEROCHOMATIN (HP1) homolog, but LHP1 performs the function similar to Pc. LHP1 associates with gene loci marked by H3K27me3 in vivo (Turck et al., 2007), which chromodomain binds H3K27me3 created by PRC2 in vitro (Zhang et al., 2007). Disruption of the chromodomain abolishes H3K27me3 recognition and mimics lhp1 null mutants (Exner et al., 2009). AtRING1 proteins are required for suppressing ectopic meristem production by repressing the misexpression of KNOX I genes (Xu and Shen, 2008). RING–finger proteins including AtRING1a/b and AtBMI1a/b are involved in repressing the vegetative–to–embryo conversion by suppressing the misexpression of stem cell–related and embryogenesis–related regulators (Chen et al., 2010). Additionally, AtBMI1c displays materially imprinting expression in endosperm (Bratzel et al., 2012). However, different from the key catalytic role of RING1B in human, all these RING–finger proteins in Arabidopsis can monoubiquitinate H2A in vitro or in vivo (Bratzel et al., 2010; 2012), indicating possibly functional divergence of PRC1 between animal and plant. Notably, DRIP1/AtBMI1a and DRIP2/AtBMI1b have been confirmed to mediate DREB2A ubiquitination in vitro, consequently act as negative regulators in drought–induced gene response by targeting DREB2A to 26S proteasomes (Qin et al., 2008).
Embryonic Flower 1 (EMF1) is thought to be a plant–specific PcG protein, no significant homolog found in other organisms (Calonje et al., 2008). Defection in EMF1 results in a broad spectrum of phenotype. Weak allele emf1–1 displays sessile cotyledons with oval shape, short hypocotyl, and lacks rosette leaves. In fact, a small inflorescence develops directly from the germinated embryo or callus; the inflorescence shoot contains sessile leaves and few sterile flowers with incomplete floral organ development, usually devoid of petals. Strong allele emf1–2 additionally displays delayed germination, and carpelloid structures formed from all lateral organs, including the cotyledons (Chen et al., 1997).
EMF1 HOMOLOGS ARE ONLY FOUND IN ANGIOSPERM
Previous reports show that EMF1 homologs are only found in plant. Here, we screened EMF1–like proteins by protein BLAST using Arabidopsis EMF1 as query against phytozome database, finding that EMF1 homologs are only exist in angiosperm, including dicot and monocot plants, not in clubmoss, brytophyte, or lower plants. Phylogenic analysis suggested that EMF1 homologs are grouped into two classes: dicot lineage and monocot lineage (Figure 1).

Before, it has been reported that Arabidopsis EMF1 contains nuclear localization signals (NLSs), ATP/GTP binding motif (P–loop), and LXXLL motif (Aubert et al., 2001). In fact, based on our multiple BLAST analyses, these putatively specific motifs are not conserved in the other most of EMF1 homologs. So, detailed domain architecture prediction in SMART database (http://smart.emblheidelberg.de/) was performed, showing there are no well–established domains existing in EMF1–like proteins. Additionally, multiple sequence alignment showed there are three highly conserved regions (CRs) with unknown function found in both dicot and monocot lineages (Figure 2).

EXPRESSION PATTERN AND SUBCELLULAR LOCALIZATION OF EMF1
RNA blot showed that EMF1 have ubiquitous and constitutive expression in all examined organs, such as root, leave, stem, and different stages of flowers (Aubert et al., 2001). Western blot using emf2–2/EMF1::EMF1–FLAG recovery transgenic line also confirmed constitutive expression of EMF1 in protein level (Calonje et al., 2008). But detailed temporal and spatial expression analysis by EMF1::GUS construct showed EMF1 promoter still has some tissue specificity, through is active in most developmental stages. The GUS activity was detected in mature embryo, in cotyledon and shoot apex in 7–day–old seedlings; in rosette leaf blade; in the 15–day–old root tips; in the stigma and anthers in opening flower, but absent in 4–day–old seedlings and floral buds excluding developing stigma papilla (Sanchez et al., 2009; Park et al., 2011). Indeed, analysis on Gene investigator also shows EMF1 have ubiquitous expression on the whole but inconsistent levels in different organs (supplemental Fig.1).
Different types of EMF1–GFP fusion constructs including full–length or truncated EMF1 versions transiently expressed in tobacco leaves showed EMF1 localized exclusively to the nucleus in a speckle–like pattern and the region between residues 337and 866 might be responsible for EMF1 subnuclear pattern (Calonje et al., 2008).
The Role of EMF1 in Different Development Phases
Maintenance of shoot apical meristem (SAM) is coordinately controlled by both KNOX pathway and WUS pathway, critical for the development of aerial parts in plant. EMF1 can directly bind the chromatin of some genes belonging to KNOX I pathway (STM, KNAT1, and KNAT2) and WUS pathway (WUS and CLV3) (Kim et al., 2012). But removal of EMF1 activity only leads to the upregulated expression of KNOX I genes, indicating that EMF1 influences the plant development mainly through directly repressing the misexpression of KNOX genes rather than WUS pathway. Overexpression of either STM or KNAT2 is able to induce ectopic carpel formation (Scofield et al., 2007), consistent with the ectopic carpelloid structure produced in emf1 mutant.
Microarray analysis showed floral time genes and floral pathway integrators, such as LFY, FT, CO and SOC1 have similar expression levels in emf1 mutant compared with WT; whereas flower organ identity genes AGAMOUS (AG), PISTILLATA (PI), APETALA1 (AP1), AP3, SEPALLATA1 (SEP1), SEP2, and SEP3 are upregulated in emf (Moon et al., 2003), consistent with the phenotype of inflorescence–to–flower transition in emf1. Indeed, LFY::GUS activity in emf mutants is similar as WT, and is confined strictly to shoot apical region, whereas AP1::GUS construct has ectopic expression in cotyledon and hypocotyl at 7DAG (Moon et al., 2003). Further ChIP analysis suggests EMF1 directly bind to the promoter and 2nd intron of AG, not the downstream region. However, EMF1 fail to bind to AG sites in the emf2–2 background, indicating EMF1 requires EMF2 to bind to AG gene and EMF1 acts downstream of EMF2. EMF1 could also be associated with the AP3 and PI promoter. But EMF1 binding to AP3 and PI loci is less influenced in emf2–2 (Calonje et al., 2008). FLC is upregulated in emf, and is also the direct target of EMF1 (Kim et al., 2009), but emf1 is extremely early flowering, indicating FLC is not the key regulator for flowering time in emf mutant. On the other hand, plenty of seed maturation genes are upregulated, moreover, embryonic regulator ABI3 and some seed storage genes are also the direct targets of EMF1 (Kim et al., 2010; 2012). So, expression level and pattern analysis suggested that EMF1 maintains vegetative development mainly by repressing the misexpression of the genes involved in flowering and embryo development.
The emf1 mutants skip vegetative growth, flowering upon germination (Sung et al., 1992), but EMF1 overexpression cannot delay flowering or induce other obvious phenotype in wild–type plants. As we known, KNAT1 promoter is active in the SAM, stronger in peripheral and rib zone than in the central zone (Lincoln et al., 1994). LFY promoter is initially active in leaf primordia at 4 day after germination (DAG) (Blazquez et al., 1997), is gradually up–regulated in leaf primordia during vegetative development, peaking in flower meristem (FM) (Nilsson et al., 1998), and also has activity in embryo, but no expression in the SAM. Seed storage protein At2S3 is expressed in embryo (Guerche et al., 1990), and in cotyledon of germinating seedling, but absent in shoot apex and rosette leaves (Sanchez et al., 2009). Tissue–specific EMF1 constructs, such as KNAT1::EMF1, LFY::EMF1, and At2S3::EMF1, failed to completely or partially recovery the emf1 phenotype. However, tissue–specific reduction of EMF1 expression in WT leads to different degrees of developmental defects. For example, SAM–specific KNAT1::asEMF1 (antisense EMF1) transgenic plants displayed obvious emf phenotype, lacking vegetative development and directly proceeding to reproductive stage upon germination, combined with no obvious phenotype in KNAT1::EMF1 line, together indicating that EMF1 activity in the SAM is essential but not sufficient to maintain vegetative development (Sanchez et al., 2009). Unexpectedly, FM–specific AP1::asEMF1 transgenic lines exhibit WT–like phenotype with normal flowering time and floral organs, indicating EMF1 is not important for flower development. So it is predicted that the defective floral organ development in emf1 might be the secondary or comprehensive effects caused by widespread release of multiple flower homeotic genes (Sanchez et al., 2009). Leaf primordial/FM–specific LFY::asEMF1 transgenic line is similar to PRC2 mutant clf, producing curly leaves, early flowering and terminal flower. LFY::asEMF1 also displays abnormal flower absent of petal. Moreover, due to EMF1 reduction in FM having no obvious effect on plant development testified by AP1::asEMF1 line, together indicating that EMF1 expression in leaf primordial has no effect on true leaf development, but has a secondary effect on flower development (Sanchez et al., 2009). Seed–specific At2S3::asEMF1 transgenic lines initially germinate as emf–like phenotype, but develop rosette leaves after 2 weeks, subsequently producing early flowering, abnormal and terminal flowers (Sanchez et al., 2009). So, reduction of EMF1 activity only in embryogenesis can greatly influence the development of next generation possibly through disturbing stable epigenetic memory deposited by EMF1. In summary, based on the analysis on expression and function aspects, EMF1 mainly maintain vegetative shoot apical meristem via directly repressing the ectopic expression of the genes related to flower development and embryogenesis
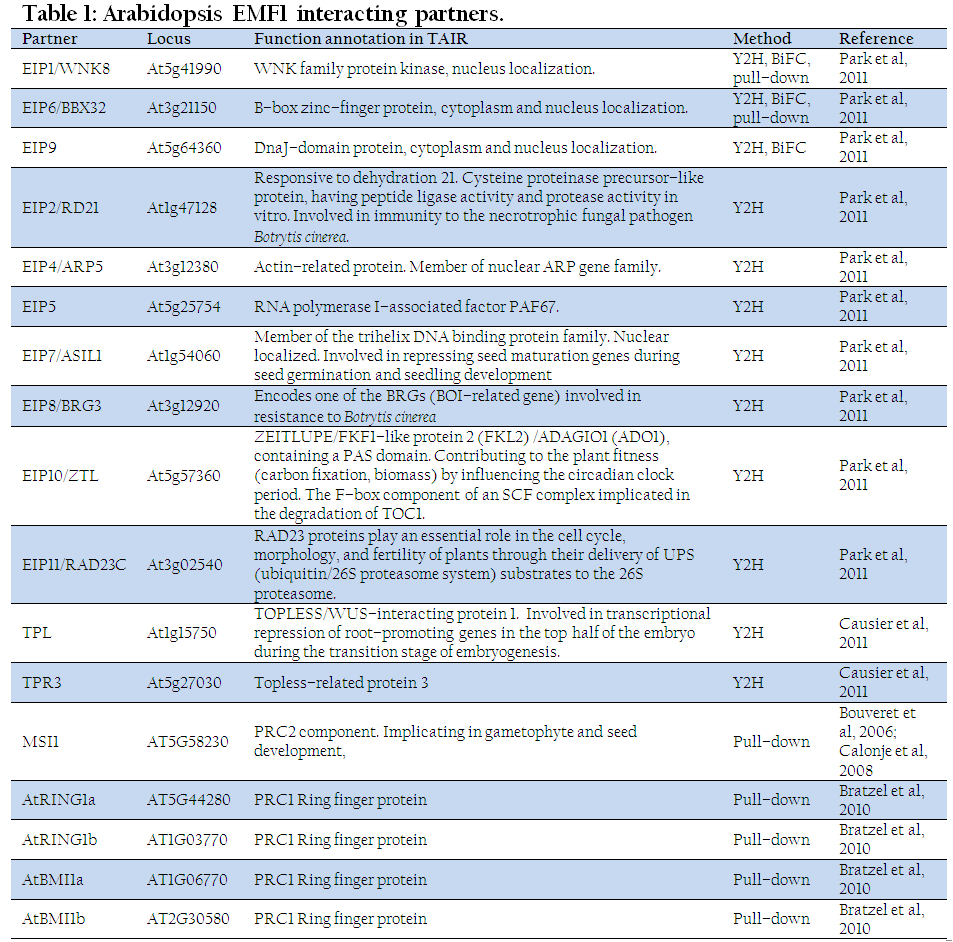
In addition, identification of EMF1 interacting proteins (EIPs) further suggested the role of EMF1 in regulating flowering time and repressing vegetative–to–embryo conversion (Table 1). Damage of each EIP1/WNK8, EIP6/BBX32, or EIP9 results in early flowering, whereas overexpression of each leads to later flowering. EIP7/ASIL1 participates in repressing seed maturation genes during vegetative development (Gao et al., 2009). Besides, EMF1 interacting factors are implicated in other pathways, such as regulation of circadian clock period (EIP10/ZTL), pathogen resistance (EIP2/RD21, EIP8/BRG3), repression of root development in shoot (TPL, TPR3), and PcG silencing (AtRING1, AtBMI1, MSI1).
EMF1 SERVES AS A NOVEL PcG-LIKE PROTEIN
EMF1 has putative PRC1 function. Firstly
Phenotype analysis of emf1;emf2 double mutant showed that emf1 is epistatic to or downstream of PRC2 component emf2 in the same pathway (Calonje et al., 2008). Secondly, EMF1 can interact with multiple PRC1 core components, such as RING–finger proteins RING1a/b, BMI1a/b. Indeed, the amount of H2Aub was reduced in emf1–2 mutants compared to WT, suggesting EMF1 is required for H2AK119 monoubiquitination activity on specific target genes (Bratzel et al., 2010). Thirdly, EMF1 can bind DNA and RNA in a non–sequence–specific fashion via its M1 (amino acids 337–617) and Ct (amino acids 866–1096) regions, and interfere with in vitro transcription fulfilled by RNA polymerase II and T7 RNA polymerase via its M1, M2 (amino acids 622–866) and Ct regions, consistent with the Drosophila PRC1 core component PSC inhibitory effect on transcription in vitro (King et al., 2002). Additionally, RING1–like proteins usually contain a conserved RING domain in N terminus and RAWUL domain in C terminus. PSC is RING1–like protein, but much larger than its homologs in human and plant. Besides both RING domain and RAWUL domain (amino acids 371–465) localized in N terminus, PSC also contains another highly charged and structurally disordered C–terminal region (PSC–CTR) (amino acids 456–1603), which binds DNA tightly and inhibit chromatin remodeling efficiently (Beh et al., 2012). Repressive PSC–CTR is absent in plant RING1 homolog, instead a putatively functional analog EMF1 which displays similarly physical properties rather than sequence similarity with PSC–CTR performs relevant function to inhibit chromatin remodeling efficiency. For instance, EMF1 homologs in both Arabidopsis thaliana and Aquilegia vulgaris exhibit largely disordered sequence with low contiguous negative charge, consistent with the criteria for repressive PSC–CTRs, showing a disordered domain with dispersed negative charges. So plant EMF1 has PSC–CTR–like properties (Beh et al., 2012).
EMF1 might be a putative PRC2 member as well. Firstly
Weak emf1 mutant phenocopies emf2, indicating both might have similar role in developmental regulation. Secondly, pull–down assay showed that EMF1 can interact with MSI1 through the region M1 (amino acids 337–617) and Ct (amino acids 866–1096), whereas no interact with other EMF2–PRC2 members, such as EMF2, CLF and FIE (Calonje et al., 2008). In fact, MSI1 is also implicated in other PRC2 complexes, like VRN2–PRC2 and FIS2–PRC2. In addition, MSI1 can interact with PRC1 member LHP1 to form a positive feedback loop to recruit PRC2 to H3K27me3–carried chromatin (Derkacheva et al., 2013). MSI1 can form a complex with RBR1 that is required for activation of the imprinted genes FIS2 and FWA (Jullien et al., 2008). Moreover, MSI1 can together with FASCIATA1 (FAS1) and FAS2 forms chromatin assembly factor 1 (CAF–1) complex (Kaya et al., 2001). But according to the mutant phenotype displayed by PRC or CAF components, EMF1 is more inclined to a PcG protein. Thirdly, EMF1 function mainly cooperates with PRC2. Among all the genes marked by H3K27me3 in WT, over 40% display reduced H3K27me3 in emf1, less than PRC2 mutants emf2 (Calonje et al., 2008; Kim et al., 2012). So, besides classical PRC2 components, EMF1 is also required for H3K27me3 deposition. EMF1 binding pattern across euchromatin covers the entire transcription unit with the peak around the transcriptional start site, displaying strong correlation with global H3K27me3 profile (Kim et al., 2012). Genes with high/low H3K27me3 deposition often exhibit high/low enrichment level of EMF1 binding. The EMF1–bound genes with high H3K27me3 are always involved in meristem, leaf, vascular, root, flower, seed development and all sorts of phytohormone response. However, The EMF1–bound genes absent of H3K27me3 markers tend to be actively transcribed genes, and always participate in cellular organization and biogenesis, cytosol and chloroplast. For instance, many photosynthesis–related genes are down–regulated in emf1 (Kim et al., 2010), suggesting EMF1 might promote their expression mainly in other indirect ways. Finally, EMF1 can counteract TrxG function. ULTRAPETALA1 (ULT1) can interact with ARABIDOPSIS TRITHORAX1 (ATX1), which belongs to trithorax group (TrxG) and catalyzes lysine–3 on histone 4 (H3K4me3), opposite to PcG with H3K27me3 activity (Saleh et al., 2007). Removal of ULT1 activity which leads to late flowering and production of extra sepals and petals can rescue the early flowering and curly leaf of PRC2 mutant clf (Carles et al., 2009). Although ult1 cannot rescue emf1 phenotype at least at very early germinating stage, it can rescue both the clf–like phenotype and most of the misregulated gene expression in leaf primordia–specific LFY::asEMF1 silencing line, consistent with the major role of ULT1 in flower development (Pu et al., 2013). So, ULT1 can antagonize EMF1 action mainly in floral transition stage, not in germinating and vegetative stage. ULT1 is upregulated in emf1 and emf2 mutants (Kim et al., 2012). Moreover, EMF1 can bind the chromatin of ULT1 and ULT2 (Pu et al., 2013), indicating EMF1 directly repressing ULT1 expression.
CONCLUSIONS
EMF genes were firstly characterized to associate with shoot maturation and development through genetic screening (Yang et al., 1997). Disruption of EMF loci results in extremely early flowering, due to absence of vegetative development. Transcriptomic analysis shows that EMF proteins maintain vegetative development mainly through suppressing the misexpression of reproductive–related genes, including flowering and seed maturation genes. Since EMF2 as a well–known PRC2 component is highly conserved in animal and plant kingdom, naturally, EMF1 has similar mutant phenotype and was thought to link to PcG function. However, EMF1 homolog only exists in plant, more precisely, in angiosperm, not found in other organisms. Certainly, animal and plant harbor so different life styles that even the homologs might evolve divergent functions, or either of them might directly evolves new components to accomplish species–specific behaviors and events. There are several important lines of evidence supporting the role of EMF1 as a PcG function. Firstly, EMF1 can produce a global influence on gene expression, consistent with epigenetic regulation. Secondly, EMF1 is required for H2Aub modification of PRC1, and EMF1 can interact with PRC1 Ring–finger proteins. Thirdly, EMF1 function cooperates with PRC2 and H3K27me3, and EMF1 can interact with PRC2 component MSI1. But there are still plenty of important questions on EMF1 need to answer, such as lacking strong evidence of in vivo protein interaction assay and further detection of precisely biochemical/regulatory function. Maybe, high throughput interactome analysis and suppressor screening can provide some better choices for deeply investigation on EMF1.
ACKNOWLEDGEMENTS
This work was supported by Research Foundation of Education Bureau of Hunan Province, China (13B045). No conflict of interest was declared.
REFERENCES
Aubert D, Chen L, Moon YH, Martin D, Castle LA, Yang CH, and Sung ZR (2001). EMF1, a novel protein involved in the control of shoot architecture and flowering in Arabidopsis. Plant Cell. 13(8):1865–1875.
http://dx.doi.org/10.2307/3871324
http://dx.doi.org/10.1105/tpc.13.8.1865
PMid:11487698 PMCid:PMC139134
Blazquez MA, Soowal LN, Lee I, and Weigel D (1997). LEAFY expression and flower initiation in Arabidopsis. Development. 124(19):3835–3844.
PMid:9367439
Beh LY, Colwell LJ, and Francis NJ (2012). A core subunit of Polycomb repressive complex 1 is broadly conserved in function but not primary sequence. Proc. Natl. Acad. Sci. U. S. A. 109(18):E1063–1071.
http://dx.doi.org/10.1073/pnas.1118678109
PMid:22517748 PMCid:PMC3344975
Bratzel F, López–Torrejón G, Koch M, Del Pozo JC, and Calonje M (2010). Keeping cell identity in Arabidopsis requires PRC1 RING–finger homologs that catalyze H2A monoubiquitination. Curr. Biol. 20(20):1853–1859.
http://dx.doi.org/10.1016/j.cub.2010.09.046
PMid:20933424
Bratzel F, Yang C, Angelova A, López–Torrejón G, Koch M, del Pozo JC, and Calonje M (2012). Regulation of the new Arabidopsis imprinted gene AtBMI1C requires the interplay of different epigenetic mechanisms. Mol. Plant. 5(1):260–269.
http://dx.doi.org/10.1093/mp/ssr078
PMid:21914649
Buchwald G, van der Stoop P, Weichenrieder O, Perrakis A, van Lohuizen M, and Sixma TK (2006). Structure and E3–ligase activity of the Ring–Ring complex of polycomb proteins Bmi1 and Ring1b. EMBO J. 25(11):2465–2474.
http://dx.doi.org/10.1038/sj.emboj.7601144
PMid:16710298 PMCid:PMC1478191
Calonje M, Sanchez R, Chen L, and Sung ZR (2008). EMBRYONIC FLOWER1 participates in polycomb group–mediated AG gene silencing in Arabidopsis. Plant Cell. 20(2):277–291.
http://dx.doi.org/10.1105/tpc.106.049957
PMid:18281509 PMCid:PMC2276442
Carles CC and Fletcher JC (2009) The SAND domain protein ULTRAPETALA1 acts as a trithorax group factor to regulate cell fate in plants. Genes Dev. 23(23): 2723–2728.
http://dx.doi.org/10.1101/gad.1812609
PMid:19952107 PMCid:PMC2788324
Causier B, Ashworth M, Guo W, and Davies B (2012). The TOPLESS interactome: a framework for gene repression in Arabidopsis. Plant Physiol. 158(1):423–438.
http://dx.doi.org/10.1104/pp.111.186999
PMid:22065421 PMCid:PMC3252085
Cernilogar FM and Orlando V (2005). Epigenome programming by Polycomb and Trithorax proteins. Biochem. Cell Biol. 83(3):322–331.
http://dx.doi.org/10.1139/o05-040
PMid:15959558
Chen D, Molitor A, Liu C, and Shen WH (2010). The Arabidopsis PRC1–like ring–finger proteins are necessary for repression of embryonic traits during vegetative growth. Cell Res. 20(12):1332–1344.
http://dx.doi.org/10.1038/cr.2010.151
PMid:21060339
Chen L, Cheng JC, Castle L, and Sung ZR (1997). EMF genes regulate Arabidopsis inflorescence development. Plant Cell. 9(11):2011–2024.
http://dx.doi.org/10.2307/3870561
http://dx.doi.org/10.1105/tpc.9.11.2011
PMid:9401124 PMCid:PMC157054
Derkacheva M, Steinbach Y, Wildhaber T, Mozgová I, Mahrez W, Nanni P, Bischof S, Gruissem W, and Hennig L (2013). Arabidopsis MSI1 connects LHP1 to PRC2 complexes. EMBO J. 32(14):2073–2085.
http://dx.doi.org/10.1038/emboj.2013.145
PMid:23778966 PMCid:PMC3715863
Exner V, Aichinger E, Shu H, Wildhaber T, Alfarano P, Caflisch A, Gruissem W, Köhler C, and Hennig L (2009). The chromodomain of LIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development. PLoS One. 4(4):e5335.
http://dx.doi.org/10.1371/journal.pone.0005335
PMid:19399177 PMCid:PMC2670505
Francis NJ, Saurin AJ, Shao Z, and Kingston RE (2001). Reconstitution of a Functional Core Polycomb Repressive Complex. Mol Cell. 8(3):545–556.
http://dx.doi.org/10.1016/S1097-2765(01)00316-1
Gao MJ, Lydiate DJ, Li X, Lui H, Gjetvaj B, Hegedus DD, and Rozwadowski K (2009). Repression of seed maturation genes by a trihelix transcriptional repressor in Arabidopsis seedlings. Plant Cell. 21(1):54–71.
http://dx.doi.org/10.1105/tpc.108.061309
PMid:19155348 PMCid:PMC2648069
Guerche P, Tire C, de Sa FG, De Clercq A, Van Montagu M, and Krebbers E (1990). Differential expression of the Arabidopsis 2S Albumin genes and the effect of increasing gene family size. Plant Cell. 2(5):469–478.
http://dx.doi.org/10.1105/tpc.2.5.469
http://dx.doi.org/10.2307/3869096
PMid:12354963 PMCid:PMC159903
Haung MD and Yang CH (1998). EMF genes interact with late–flowering genes to regulate Arabidopsis shoot development. Plant Cell Physiol. 39(4):382–393.
http://dx.doi.org/10.1093/oxfordjournals.pcp.a029381
PMid:9615462
Hennig L and Derkacheva M (2009). Diversity of Polycomb group complexes in plants: same rules, different players? Trends Genet. 25(9):414–423.
http://dx.doi.org/10.1016/j.tig.2009.07.002
PMid:19716619
Hennig L, Taranto P,Walser M, Schonrock N, and Gruissem W (2003). Arabidopsis MSI1 is required for epigenetic maintenance of reproductive development. Development. 130(12):2555–2565.
http://dx.doi.org/10.1242/dev.00470
PMid:12736201
Jürgens G (1985). A group of genes controlling the spatial expression of the bithorax complex in Drosophila. Nature. 316(6024):153–155.
http://dx.doi.org/10.1038/316153a0
Jullien PE, Mosquna A, Ingouff M, Sakata T, Ohad N, and Berger F (2008). Retinoblastoma and its binding partner MSI1 control imprinting in Arabidopsis. PLoS Biol. 6(8):e194.
http://dx.doi.org/10.1371/journal.pbio.0060194
PMid:18700816 PMCid:PMC2504488
Kaya H, Shibahara KI, Taoka KI, Iwabuchi M, Stillman B, and Araki T (2001). FASCIATA genes for chromatin assembly factor–1 in arabidopsis maintain the cellular organization of apical meristems. Cell. 104(1):131–142.
http://dx.doi.org/10.1016/S0092-8674(01)00197-0
Kim SY, Lee J, Eshed–Williams L, Zilberman D, and Sung ZR (2012). EMF1 and PRC2 cooperate to repress key regulators of Arabidopsis development. PLoS Genet. 8(3):e1002512.
http://dx.doi.org/10.1371/journal.pgen.1002512
PMid:22457632 PMCid:PMC3310727
Kim SY, Zhu T, and Sung ZR (2010). Epigenetic regulation of gene programs by EMF1 and EMF2 in Arabidopsis. Plant Physiol. 152(2):516–528.
http://dx.doi.org/10.1104/pp.109.143495
PMid:19783648 PMCid:PMC2815887
Lewis EB (1978). A gene complex controlling segmentation in Drosophila. Nature. 276(5688): 565–570.
http://dx.doi.org/10.1038/276565a0
PMid:103000
Lincoln C, Long J, Yamaguchi K, Serikawa K, and Hake S (1994). A Knotted1–like homeobox gene in Arabidopsis is expressed in the vegetative meristem and dramatically alters leaf morphology when overexpressed in transgenic plants. Plant Cell. 6(12):1859–1876.
http://dx.doi.org/10.1105/tpc.6.12.1859
http://dx.doi.org/10.2307/3869913
PMid:7866029 PMCid:PMC160567
Lund AH and van Lohuizen M (2004). Polycomb complexes and silencing mechanisms. Curr. Opin. Cell Biol. 16(3):239–246.
http://dx.doi.org/10.1016/j.ceb.2004.03.010
PMid:15145347
Molitor A and Shen WH (2013). The polycomb complex PRC1: composition and function in plants. J. Genet. Genomics. 40(5):231–238.
http://dx.doi.org/10.1016/j.jgg.2012.12.005
PMid:23706298
Moon YH, Chen L, Pan RL, Chang HS, Zhu T, Maffeo DM, and Sung ZR (2003). EMF genes maintain vegetative development by repressing the flower program in Arabidopsis. Plant Cell. 15(3):681–693.
http://dx.doi.org/10.1105/tpc.007831
PMid:12615941 PMCid:PMC150022
Nilsson O, Wu E, Wolfe D, and Weigel D (1998). Genetic ablation of flowers in transgenic Arabidopsis. Plant J. 15(6):799–804.
http://dx.doi.org/10.1046/j.1365-313X.1998.00260.x
PMid:9807818
Park HY, Lee SY, Seok HY, Kim SH, Sung ZR, and Moon YH (2011). EMF1 interacts with EIP1, EIP6 or EIP9 involved in the regulation of flowering time in Arabidopsis. Plant Cell Physiol. 52(8):1376–1388.
http://dx.doi.org/10.1093/pcp/pcr084
PMid:21700722
Pien S and Grossniklaus U (2007). Polycomb group and trithorax group proteins in Arabidopsis. Biochem Biophys Acta. 1769(5-6):375-382.
http://dx.doi.org/10.1016/j.bbaexp.2007.01.010
PMid:17363079
Pu L, Liu MS, Kim SY, Chen LF, Fletcher JC, and Sung ZR (2013). EMBRYONIC FLOWER1 and ULTRAPETALA1 act antagonistically on Arabidopsis development and stress response. Plant Physiol. 162(2):812–830.
http://dx.doi.org/10.1104/pp.112.213223
PMid:23632855 PMCid:PMC3668072
Qin F, Sakuma Y, Tran LS, Maruyama K, Kidokoro S, Fujita Y, Fujita M, Umezawa T, Sawano Y, Miyazono K, Tanokura M, Shinozaki K, and Yamaguchi–Shinozaki K (2008). Arabidopsis DREB2A–interacting proteins function as RING E3 ligases and negatively regulate plant drought stress–responsive gene expression. Plant Cell. 20(6):1693–1707.
http://dx.doi.org/10.1105/tpc.107.057380
PMid:18552202 PMCid:PMC2483357
Saleh A, Al–Abdallat A, Ndamukong I, Alvarez–Venegas R, and Avramova Z (2007). The Arabidopsis homologs of trithorax (ATX1) and enhancer of zeste (CLF) establish 'bivalent chromatin marks' at the silent AGAMOUS locus. Nucleic Acids Res. 35(18): 6290–6296.
http://dx.doi.org/10.1093/nar/gkm464
PMid:17881378 PMCid:PMC2094062
Sánchez R, Kim MY, Calonje M, Moon YH, and Sung ZR (2009). Temporal and spatial requirement of EMF1 activity for Arabidopsis vegetative and reproductive development. Mol. Plant. 2(4):643–653.
http://dx.doi.org/10.1093/mp/ssp004
PMid:19825645
Sanchez–Pulido L, Devos D, Sung ZR, and Calonje M (2008). RAWUL: a new ubiquitin–like domain in PRC1 ring finger proteins that unveils putative plant and worm PRC1 orthologs. BMC Genomics. 9:308.
http://dx.doi.org/10.1186/1471-2164-9-308
PMid:18588675 PMCid:PMC2447854
Scofield S, Dewitte W, and Murray JA (2007). The KNOX gene SHOOT MERISTEMLESS is required for the development of reproductive meristematic tissues in Arabidopsis. Plant J. 50(5):767–781.
http://dx.doi.org/10.1111/j.1365-313X.2007.03095.x
PMid:17461793
Shao Z, Raible F, Mollaaghababa R, Guyon JR, Wu CT, Bender W, and Kingston RE (1999). Stabilization of Chromatin Structure by PRC1, a Polycomb Complex. Cell. 98(1): 37–46.
http://dx.doi.org/10.1016/S0092-8674(00)80604-2
Sung ZR, Belachew A, Shunong B, and Bertrand–Garcia R (1992). EMF, an Arabidopsis gene required for vegetative shoot development. Science. 258(5088):1645–1647.
http://dx.doi.org/10.1126/science.258.5088.1645
PMid:17742537
Tamura K, Dudley J, Nei M, and Kumar S (2007). MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24(8):1596–1599.
http://dx.doi.org/10.1093/molbev/msm092
PMid:17488738
Thompson JD, Higgins DG, and Gibson TJ (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position–specific gap penalties and weight matrix choice. Nucleic Acids Res. 22(22):4673–4680.
http://dx.doi.org/10.1093/nar/22.22.4673
PMid:7984417 PMCid:PMC308517
Turck F, Roudier F, Farrona S, Martin–Magniette ML, Guillaume E, Buisine N, Gagnot S, Martienssen RA, Coupland G, and Colot V (2007). Arabidopsis TFL2/LHP1 specifically associates with genes marked by trimethylation of histone H3 lysine 27. PLoS Genet. 3(6):e86.
http://dx.doi.org/10.1371/journal.pgen.0030086
PMid:17542647 PMCid:PMC1885283
Wang H, Wang L, Erdjument–Bromage H, Vidal M, Tempst P, Jones RS, and Zhang Y (2004). Role of histone H2A ubiquitination in Polycomb silencing. Nature. 431(7010):873–878.
http://dx.doi.org/10.1038/nature02985
PMid:15386022
Xu L and Shen WH (2008). Polycomb silencing of KNOX genes confines shoot stem cell niches in Arabidopsis. Curr. Biol. 18(24):1966–1971.
http://dx.doi.org/10.1016/j.cub.2008.11.019
PMid:19097900
Yang CH, Chen LJ, and Sung ZR (1995). Genetic regulation of shoot development in Arabidopsis: role of the EMF genes. Dev. Biol. 169(2):421–435.
http://dx.doi.org/10.1006/dbio.1995.1158
PMid:7781888
Zhang X, Germann S, Blus BJ, Khorasanizadeh S, Gaudin V, and Jacobsen SE (2007). The Arabidopsis LHP1 protein colocalizes with histone H3 Lys27 trimethylation. Nat. Struct. Mol. Biol. 14(9):869–871.
http://dx.doi.org/10.1038/nsmb1283
PMid:17676062

