
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 1 (4): 116 – 119β–Glucan from Edible Mushroom (Pleurotus florida) Enhances Mucosal Immunity in Poultry
Gopi Muthusamy1, Siddhartha Narayan Joardar1*, Indranil Samanta1, Devi Prasad Isore1, Barun Roy2, Tapas Kumar Maiti3
- Department of Veterinary Microbiology, Faculty of Veterinary and Animal Sciences, West Bengal University of Animal and Fishery Sciences, Kolkata–700037
- Department of Animal Nutrition, Faculty of Veterinary and Animal Sciences, West Bengal University of Animal and Fishery Sciences, Kolkata–700037
- Department of Biotechnology, Indian Institute of Technology, Kharagpur–721302
*Corresponding author:[email protected],
ARTICLE CITATION:
Muthusamy G, Joardar SN, Samanta I, Isore DP, Roy B and Maiti KT (2013). β–Glucan from edible mushroom (Pleurotus florida) enhances mucosal immunity in poultry. Adv. Anim. Vet. Sci. 1 (4): 116 – 119.
Received: 2013–08–06, Revised: 2013–08–17, Accepted: 2013–08–18
The electronic version of this article is the complete one and can be found online at
(
http://www.nexusacademicpublishers.com/table_contents_detail/4/81/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
The study was undertaken to assess the effect of orally administered β–glucan on intestinal intra epithelial leukocytes (iIEL) of broiler to establish its role in mucosal immunity. After acclimatization for 7 days with control diet, two groups of broiler birds each containing 20 chicks, with replica of 10 chicks in each pen, were fed with experimental feed containing β–glucan @15mg/ kg (group A) and 30 mg / kg (group B) for a period of 20 days and, then given normal diet. Birds kept with normal feeding regime (n=10) were considered as control (group C). Intestinal intraepithelial leucocytes (iIEL) of the experimental birds were isolated and in–vitro immune–effectors activities were observed. Significant enhancement (P ≤ 0.05) of In vitro phagocytic activity, nitrite production, proliferative activity and cytokine production of iIEL were observed in birds of group B than C. The results showed that supplementation of β–glucan from edible mushroom in feed (30mg/ kg) for 20 days had significant immunostimulatory effect in broiler birds that was reflected in the In vitro immune–effector activities of the mucosal cells (iIEL).
INTRODUCTION
Infectious diseases are the major challenge in commercial poultry industry. Preventive health management through vaccination is universally recommended practice to meet this challenge, although, cases of vaccine failure and disease outbreak are also common (Dhama et al., 2013).
Immunostimulants comprise of a group of biological and synthetic compounds that enhance the non–specific cellular and humoral defense mechanisms in animals (Chan et al, 2009). It also increases resistance to infectious diseases, and use of these immunostimulants is an effective means of increasing the immunocompetence and disease resistance of animals, birds and fishes. Generally, immunostimulants are promoting a greater and more effective sustained immune response to the infectious agent without the risks of toxicity, carcinogenicity or tissue residues (Weickert and Pfeiffer, 2008; Firenzouli et al., 2008). Beta glucans from the yeast cell wall and mushroom have been shown to stimulate both specific and non–specific immune responses and improve the growth performance also (Vetvicka et al., 2008). However, it has been observed that glucans from various sources appear to elicit diverse immunomodulatory effects in numerous animal tissues, including the blood, gastrointestinal tract and spleen (Ramberg et al., 2010; Samulsen et al., 2011). The use of immunostimulants that is β–1, 3/1–6–glucan can protect Staphylococcus aureus, Listeria monocytogens, Bacillus anthracis, Venezuelan equine encephalomyelitis infection in mice, Salmonella enterica serovar Enteritidis infection in chicken and Staphylococcus aureus infection in cattle (Li et al., 2004; Lowry et al., 2005; Tominac et al., 2010).
Immunostimulants should be active by oral route and should be stable both in its native state and after incorporation into food and water. Oral administrations of immunostimulants have already been reported for glucans, lactoferrin, levamisole and chitosan (Kamilya et al., 2008). Oral administration is the most practical method for delivery of immunostimulants and results in enhancement of leucocyte function and protection against infectious diseases (Tominac et al., 2010).
Studies on immunomodulatory role of edible mushroom Pleurotus florida in fish has produced encouraging results. It was observed that extracted proteoglycon/glucan from Pleurotus florida when used in feed enhanced the activities of immune effector cells of fish and the protection against pathogen (Aeromonas hydrophila) challenge in experimental condition (Kamilya et al., 2008). In vitro immunostimulatory effects as also adjuvancity of mushroom glucan was tested in fish (Kamilya et al., 2006). Earlier, we have shown that orally administered β–glucan of edible mushroom (Pleurotus florida) up–regulates innate immune response in broiler (Paul et al., 2012). Modulation of immune–effector activities of haematopoietic cells viz. neutrophil and blood mononuclear cells were reported in that study. Here we report, the effect of orally administered β–glucan of edible mushroom (Pleurotus florida) on intestinal intra epithelial leukocytes (iIEL) of broiler to establish its role in mucosal immunity.
MATERIALS AND METHODS
Immunostimulant
Purified Mushroom Glucan (MG) was prepared from the source of Ostreus Mushroom by alkali extraction method (Maiti et al., 2008). The purified product was obtained from the Department of Biotechnology, Indian Institute of Technology, Kharagpur, India.
Experimental Bird
The experimental design and procedure adopted was approved by the Animal Ethics Committee of the University. Fifty day– old chicks were procured from a commercial hatchery and vaccinated at 5th day with RD vaccine (F strain) (Institute of Animal Health and Veterinary Biologicals, Govt. of West Bengal). On 7th day, they were divided into three diet groups (Group–A, B, C). A and B group contained 20 chick with replica of 10 chicks in each pen. Control group (C) had 10 birds with replica of 5 chicks in each pen.
Experimental Design
Birds were fed (normal) feed (Amrit Feed, India) for 20 days. On 7th day onward, the experimental birds of group A were given 15 mg MG / kg whereas, birds in the Group B were given 30 mg / kg in feed. Group C birds (Control) were given normal feed. After 20 days of feeding, birds of all three groups (A, B, C) were given normal feed for further 14 days. Birds were allowed to take feed and water ad libitum during the whole period of experiment. The period of observation was started from the last MG administration (0–day) with 7 days interval, viz. 0 day, 7th day and 14th day.
Isolation of intestinal Intra Epithelial leukocyte cells (iIEL)
Three birds of each group (viz. Group A, Group B and Group C) were sacrificed under anesthesia (following the standard guidelines of Institutional Ethical Committee) to collect the intestine in sterile PBS on each experimental day. The iIEL were isolated from chicken intestine with slight modification as adapted by Grachia et al. (1997). In brief, 12 to 15 cm of duodenal C loops, jejunum, ileum and caeca were removed from chicken intestine, washed with ice cold PBS buffer with antibiotics solution (containing streptopenicillin @400 IU/mL, gentamycin @4mg/mL) extensively. The gut tissues were taken into a beaker treated with pre warmed (41ºC) 5mM dithiothreitol (DTT) and 0.1 mM Ethylene Diamine Tetra Acetic acid (EDTA) solution for 40 minutes in water bath (temperature 41ºC) with occasional gentle shaking. After extensive washing, the treated intestinal tissues were placed in a beaker containing 30 ml of washing medium having 300 IU of collagenase per mL and kept in a shaking water bath (41ºC). After 30 min, supernatants containing single cells were collected and replaced with fresh washing medium containing collagenase and incubated for an additional 30 min at 41°C. The viable cells (iIEL) were collected by centrifugation, washed twice in washing medium and separated from debris and dead cells by differential centrifugation using HistopaqueR (Sigma, USA). Viability was assessed by trypan blue dye exclusion method (Daly et al., 1995). Then the iIEL cells were used for the functional assays.
Stock solutions for LPS and ConA
Stock solution of lipopolysaccharide (Sigma, USA) was prepared at a concentration of 20μg/mL of the growth medium RPMI–1640 (Sigma, USA), filtered through a sterile membrane filter (0.2μ) and stored at –20ºC until use. Stock solution of concanavalin A (Sigma, USA) was prepared at the concentration of 20μg/mL of the growth medium RPMI–1640 (Sigma, USA), filtered through a sterile membrane filter (0.2μ) and stored at –20ºC until use.
In vitro Nitrite Production Assay
The production of reactive nitrogen intermediates was assessed following the method described by Tafalla and Novoa (2000). This method was based on the Griess reaction that quantified the nitrite content of the macrophage supernatants, as nitric oxide is an unstable molecule and degrades to nitrite and nitrate (Green et al., 1982).
Lymphoproliferation Assay (LPA)
The colorimetric – [4, 5– dimethylthiazol–2–yl]–2, 5– diphenyl tetrazolium bromide (MTT) assay described by Daly et al. (1995) was used to determine the proliferation of iIEL cells.
Macrophage Functional Assay
Phagocytosis was examined as described by Yoshida et al. (1993).
cDNA Preparation
After separation from intestine, iIEL cells (2×106cells / mL) were suspended in RPMI–1640 and 100 μL of cell suspension was dispended into wells of 96–well tissue culture plates. The final volume of the wells made up to 200 μL with Con A at a concentration of 10μg/mL in positive control wells (in triplicate) and with RPMI–1640 growth medium in negative control wells (in triplicate). Finally, it was incubated at 37ºC for 3 hr containing 5% CO2 tension.
For cell harvesting, cells were thoroughly mixed with the culture medium and the cell suspension was transferred to a 15 mL polypropylene tube. The tube was centrifuged at 400g for 5 min to pellet the cells. The cells (pellet) were washed twice with 10 mL of PBS. Finally, the pellet was suspended in 1mL of culture medium. The cell density was determined and it was adjusted using ice–cold PBS so that it falls within the range of 5000 cell/µL. Now 10µL of cells were transferred to PCR tube and cDNA was prepared following the protocol mentioned in cDNA kit (GeNei™).
Detection of cytokine mRNA by RT PCR
The presence of mRNA for chicken IFN–γ was determined by RT–PCR as described earlier following the touch–down PCR protocol for amplification of cDNA (Xing and Schat, 2000). The PCR product (485 bp) was analyzed by agarose gel electrophoresis and documented in the gel documentation system (UVP). As well as the image was analyzed by UVP software (Doc ILT) for semi quantification of the cDNA present in the product.
Statistical Analysis
The results of each experiment was expressed as the mean ± standard error of mean and analyzed by one–way analysis of variance (ANOVA) to test the significance between control and experimental groups.
RESULTS
In vitro Nitrite Production Assay
Upon LPS stimulation the In vitro nitrite production of intestinal intra–epithelial leukocytes (iIEL) of broiler birds of group A, B, C are shown in Figure 1. In vitro nitrite production was increased significantly (P ≤ 0.05) in treated birds of both the groups subsequently through 0 day, 7th day and 14th day than control group. The average values of Group B birds were more than Group A in all the post treatment periods.
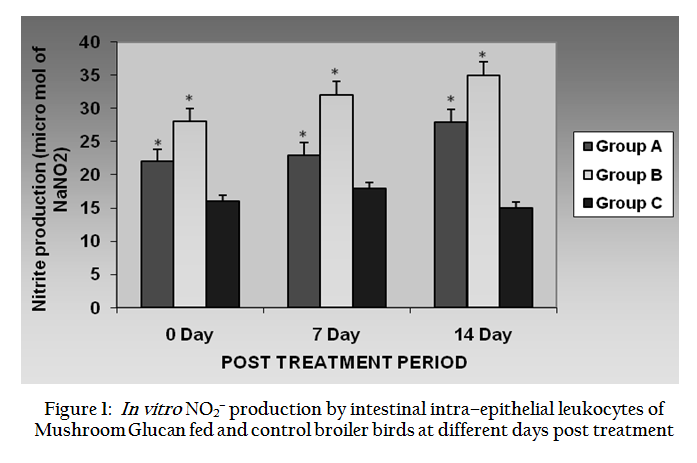
Figure 1: In vitro NO2– production by intestinal intra–epithelial leukocytes of Mushroom Glucan fed and control broiler birds at different days post treatment
Lympho–proliferation assay
Upon ConA stimulation, the In vitro iIEL of broiler birds, in group A, B and C were expressed as stimulation index (S.I.). The S.I. values of treated birds of both the groups enhanced in post treatment period, viz. 0–day, 7th day and 14th day. The average S.I. values were gradually increased from 0.157 ± 0.003 to 0.302 ± 0.001 in case of group A and 0.174 ± 0.005 to 0.331 ± 0.056 in case of group B from 0–day to 14th day. In all the cases values were more in case of Group B than Group A. These values were significantly (P ≤ 0.05) higher than that of group C in all the cases. The results are shown in the Figure 2.
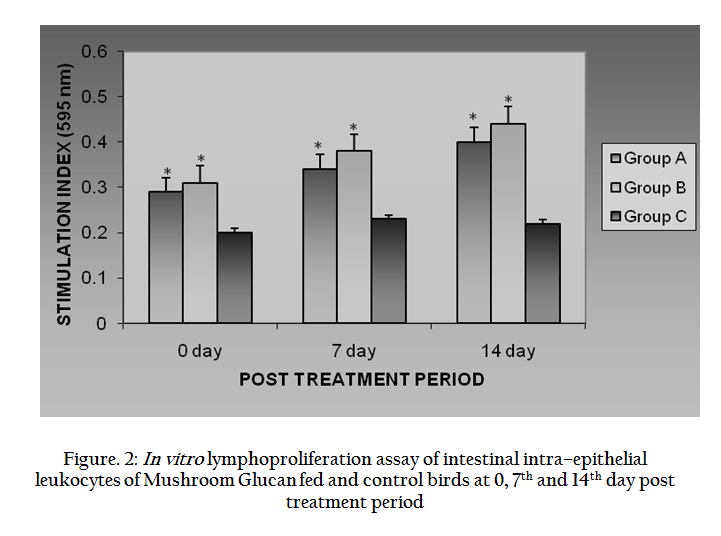
Figure 2: In vitro lymphoproliferation assay of intestinal intra–epithelial leukocytes of Mushroom Glucan fed and control birds at 0, 7th and 14th day post treatment
Macrophage functional assay
The Phagocytic Activity (PA) value of iIEL cells of control and MG fed birds are presented in Figure 3. The PA values of treated birds of both the groups enhanced significantly (P ≤ 0.05) in post treatment period, viz. 0–day, 7th day and 14th day. The average values of Group B birds were more than Group A in all the post treatment periods.
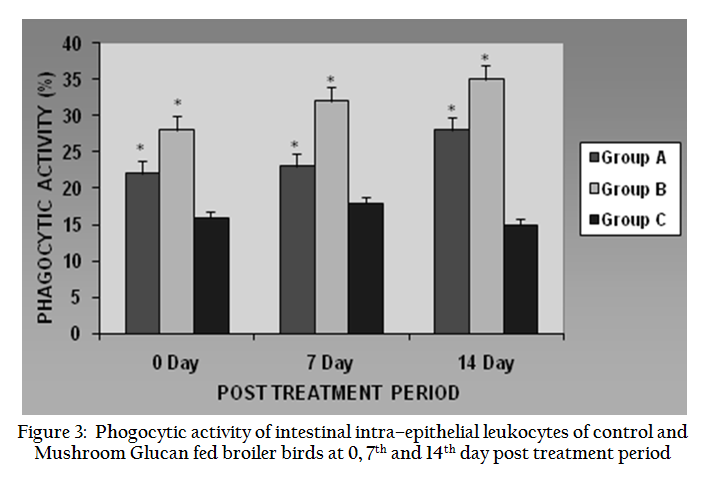
Figure 3: Phogocytic activity of intestinal intra–epithelial leukocytes of control and Mushroom Glucan fed broiler birds at 0, 7th and 14th day post treatment period
Cytokine Assay
iIEL from control and treated birds (group A and group B) were separated and stimulated by ConA In vitro. In culture, mRNA produced was reverse transcribed and amplified by RT–PCR to assess cytokines produced viz. IFN–γ using specific primers. The RT–PCR showed m–RNA production of IFN–γ In vitro by iIEL of both control and treated group birds. Birds of control groups produced less IFN–γ–cDNA in compared to birds of treated groups as per the results observed in the UVP software analysis.
DISCUSSION
This work illustrates the influence of dietary supplementation of β– glucan from edible mushroom (Pleurotus florida) on the activities of immune effector cells of broiler birds. As the transmission of various infectious diseases to animals is taken place through the alimentary tract route, understanding the nature and functions of various cellular components of gut associated lymphoid tissue (GALT) gives us a panoramic view about the type of immunological responses operated in gastrointestinal tract (GIT). Lamina propria and intestinal intra epithelial leukocytes (iIEL) are the major components of gut immunity. The iIEL are located at basal and epical part of the intestinal epithelium, which makes them virtually most important in immunosurveillance mechanism (Lee et al., 2010).
Reactive nitrogen intermediates have been proved to be important molecules in regulating immune functions, as well as having direct anti–microbial effect (Liew, 1995).
In the present study, the iIEL of MG fed broiler chicks induced by LPS In vitro was assessed for nitrite production. In all the post treatment period, treated birds showed higher In vitro nitrite production than the control birds. It is known that LPS are capable of stimulating macrophage nitrite production in mice (Stuehr and Marletta, 1985) and in fish (Tafalla and Novoa, 2000). Our finding is supported by earlier works in mammals and birds where MG augmented nitric oxide production (Mucksova et al., 2001; Cox et al., 2010). Earlier workers also showed immunostimulatory effect of β– glucan of edible mushroom origin in other animal species like Indian major carp, Catla catla (Kamilya et al., 2006). In an experiment, broiler birds fed with 20 mg β– glucan gave higher In vitro nitrite production by blood leukocytes than control birds (Paul, 2008).
There was a significant enhancing effect of dietary MG on the proliferative response of iIEL, induced by Con A. In vitro lymphoproliferation was enhanced on group A and group B birds and the stimulation index (SI) was significantly (P ≤ 0.05) more than control group in all the post treatment periods. It was found to be that administration of oat β– glucan increased lymphocyte and neutrophil stimulation in beef steers (Estrada et al., 1999). L–Arginine was observed to stimulate in virto lymphoproliferation of peripheral blood lymphocytes, iIEL and increase the systemic immune response in chickens that gave 100% protection in IBD virus challenge (Tayade et al., 2006). β– glucan of yeast cell wall origin (Saccharomyces cerevisiae) was found to have immunostimulatory effect in fish. Enhancing effect of dietary glucan on the response of lymphocytes of fish induced by ConA has been observed previously (Verlhac et al., 1998). Similarly, increased speenocyte proliferation in response to LPS has been observed following oral administration of direct fed microbials in broiler birds (Lee et al., 2010).
An enhanced phagocytic activity (PA) of iIEL was observed in MG fed birds. Group A and group B birds showed high PA value than the control group in all the post experimental periods. Earlier workers reported similar observation in other species (mice). Sakurai et al. (1992) showed enhanced murine alveolar macrophage activity due to β– glucan feeding. Suzuki et al. (1990) reported that orally administered β–glucan enhanced the phagocytic activity of macrophage in mice. Similar results were observed in chicken treated with purified β– glucan. The PA activity increased in immature chickens against Salmonella enterica serovar Enteritidis (Lowry et al., 2005).
In the present study, semi quantitative RT–PCR for mRNA was conducted to assess the In vitro production of IFN–γ from iIEL of treated and control birds stimulated by ConA. Birds of control groups produced In vitro cytokines viz. IFN–γ less as compared to birds of treated groups. This shows that leukocytes of birds from treated groups where highly sensitized than the leukocytes of control birds. That means MG had positive response in treated birds in terms of cytokine production by iIEL. Earlier worker (Rice et al., 2005) also obtained similar positive correlation between immunostimulation and cytokine production. In that study glucan as immunostimulatory carbohydrates has increased systemic levels of IL–12. The orally administered yeast β– 1, 3 glucan to mice stimulated the production of cytokines (Rice et al., 2005). However, in contrast, Cox et al. (2010) observed reduced expression of IFN–γ in broiler birds fed with beta glucan.
In the mammalian system, action of β–glucan is mediated through toll–like receptors (TLR) and dectin–1 (Brown and Gordon, 2003). It is known that after binding with dectin–1, β–glucan activates the transcription factor NF–κB that plays a critical role in immune responses (Tada et al., 2009; Samuelsen et al., 2011). The authors hypothesize such type of molecular interactions at cellular level in poultry gut that substantiates the present findings.
CONCLUSION
The results showed that supplementation of β–glucan from edible mushroom in feed (30mg/ kg) for 20 days had significant immunostimulatory effect in broiler birds as reflected in the immune–effector activities of mucosal cells (iIEL).
ACKNOWLEDGEMENT
Authors are thankful to the Vice Chancellor, WBUAFS for providing necessary research facilities.
REFERENCES
Chan GC, Chan WK and Sze DM (2009). The effects of beta–glucan on human immune and cancer cells. J. Hematol. Oncol. 2: 5–7.
http://dx.doi.org/10.1186/1756-8722-2-25
PMid:19515245 PMCid:PMC2704234
Cox CM, Summers LH, Kim S, McElroy AP, Bedford MR, Dalloul RA (2010). Immune responses to dietary beta glucan in broiler chicks during an Eimeria challenge. Poultry Sci. 89: 2597–2607.
http://dx.doi.org/10.3382/ps.2010-00987
http://dx.doi.org/10.3382/ps.2010-00865
Daly JG, Olivier G and Moore AR (1995). A calorimetric assay for the quantification of brook trout (Salvelinus fontinalis) lymphocyte mitogenesis. Fish Shellfish Immunol. 5: 266–273.
http://dx.doi.org/10.1006/fsim.1995.0026
Dhama K, Wani MY, Deb R, Karthik K, Tiwari R, Barathidasan R, Kumar A, Mahima, Verma AK, Singh SD (2013). Plant based oral vaccines for human and animal pathogens – a new era of prophylaxis: current and future prospective. J. Exp. Biol. Agri. Sci. 1(1): 1–12.
Estrada A, Van Kessel A and Laarveld B (1999). Effect of administration of oat–glucan on immune parameters of healthy and immunosuppressed beef steers. Canadian J. Vet. Res. 63: 261–268.
PMid:10534005 PMCid:PMC1189562
Firenzouli F, Gori L and Lombardo G (2008). The medicinal mushroom Agaricus blazei Murill: Review of literature and pharmaco–toxicological problems. Evid. Based complement Alternat. Med. 5:3–15.
http://dx.doi.org/10.1093/ecam/nem007
PMid:18317543 PMCid:PMC2249742
Grachia, F.J.S., Aller, W.W. and Mccormack, W.T. (1997). Impaired calcium mobilization and different tyrosine phosphorylation in intestinal intraepithelial lymphocytes. Immunol., 91: 81–87.
http://dx.doi.org/10.1046/j.1365-2567.1997.00228.x
Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS and Tannnbaum S (1982). Analysis of nitrate, nitrite and (15N) nitrate in biological fluids. Analytical Biochem. 126:131–138.
http://dx.doi.org/10.1016/0003-2697(82)90118-X
Kamilya D, Joardar SN, Mal CB and Maiti TK (2008). Effects of a glucan from the edible mushroom (Pleurotus florida) as an immunostimulant in farmed Indian Major carp (Catla catla). The Israeli J. Aquacult. 60:37–45.
Kamilya D, Maiti TK, Joardar SN and Mal BC (2006). Adjuvant effect of mushroom glucan and bovine lactoferrin upon Aeromonas hydrophila vaccination in catla, catla catla (Hamilton). J. Fish Dis. 29: 331–337.
http://dx.doi.org/10.1111/j.1365-2761.2006.00722.x
PMid:16768713
Lee KW, Lee SH, Lillehoj HS, Li GX, Jang SI, Babu US, Park MS, Kim DK, Lillehoj EP, Neumann AP, Rehberger TG and Siragusa GR (2010). Effects of direct fed microbials on growth performance, gut morphometry and immune characteristics in broiler chickens. Poultry Sci. 89: 203–216.
http://dx.doi.org/10.3382/ps.2009-00418
PMid:20075271
Liew FY (1995). Interaction between cytokines and nitric oxide. Adv. Neuroimmunol. 5: 201–209.
http://dx.doi.org/10.1016/0960-5428(95)00009-Q
Li W, Yajima T, Saito K, Nishimura H, Fushimi T, Ohshima Y, Tsukamota Y and Yoshikai Y (2004). Immunostimulating properties of intragastrically administered Acetobacter–derived soluble branched (1,4)–beta–D–glucans decrease murine susceptibility to Listeria monocytogenes. Infect. Immun. 72: 7005–7011.
http://dx.doi.org/10.1128/IAI.72.12.7005-7011.2004
PMid:15557623 PMCid:PMC529118
Lowry VK, Farnell MB, Ferro PJ, Swaggerty CL, Bahl A and Kogut MH (2005). Purified beta–glucan as an abiotic feed additive up–regulates the innate immune response in immature chickens against Salmonella enterica serovar Enteritidis. Int J. Food Microbiol. 98: 309–318.
http://dx.doi.org/10.1016/j.ijfoodmicro.2004.06.008
PMid:15698692
Maiti S, Bhutia SK, Mallick SK, Kumar A, Khadgi N and Maiti TK (2008). Antiproliferative and immunostimulatory protein fraction from edible mushrooms. Environ. Toxicol. Pharmacol. 26:187–191.
http://dx.doi.org/10.1016/j.etap.2008.03.009
PMid:21783909
Mucksova J, Babicek K and Pospisil M (2001). Particulate 1,3 beta–D–Glucan, carboxymethylglucan and sulfoethylglucan–influence of their oral or intraperitoneal administration on immunological respondence of mice. Folia Microbiol (Praha) 46 (6): 559–563.
http://dx.doi.org/10.1007/BF02818003
Pal D, Joardar SN and Roy B (2007). Immunostimulatory effects of a yeast (Saccharomyces cerevisiae) cell wall feed supplement on Rohu (Labeo rohita), an Indian Major Carp. The Israeli J. Aquacult. 59:175–182.
Paul I (2008). Immunomodulatory role of β glucan from yeast cell wall and edible mushroom origin with special reference to Ranikhet Disease. M.V.Sc thesis submitted to West Bengal University of Animal and fishery Sciences, Kolkata, India.
Ramberg JE, Nelson ED and Sinnot RA (2010). Immunomodulatory dietary polysaccharides: A systematic review of the literature. Nutrition J. 9: 54–76.
http://dx.doi.org/10.1186/1475-2891-9-54
PMid:21087484 PMCid:PMC2998446
Rice P J, Adams EL, Ozment–Skelton T, Gonzalez AJ, Goldman MP and Lockhart BE (2005). Oral delivery and gastrointestinal absorption of soluble glucans stimulate increased resistance to infectious challenge. The J. Pharmacol. Exp. Therapeutics 314: 1079–1086.
http://dx.doi.org/10.1124/jpet.105.085415
PMid:15976018
Samulsen AB, Rieder A, Grimmer S, Michaelsen TE and Knutsen S (2011). Int. J. Mol. Sci. 12:570–587.
http://dx.doi.org/10.3390/ijms12010570
PMid:21340001 PMCid:PMC3039967
Sakurai T, Hashimoto K, Suzuki I, Ohno N, Oikawa S and Masuda A (1992). Enhancement of murine alveolar macrophage functions by orally administered beta–glucan. Inter J. Immunopharmacol. 14: 821–830.
http://dx.doi.org/10.1016/0192-0561(92)90080-5
Stuehr DJ and Marletta MA (1985). Mammalian nitrate biosynthesis: mouse macrophages produce nitrite and nitrate in response to Escherichia coli lipopolysaccharide. Proc. Natl. Acad. Sci. USA 82: 7738–7742.
http://dx.doi.org/10.1073/pnas.82.22.7738
PMid:3906650 PMCid:PMC391409
Suzuki I, Tanaka H, Kinoshita A, Oikawa S, Osawa M and Yadomae T (1990). Effect of orally administered beta–glucan on macrophage function in mice. Inter. J. Immunopharmacol. 12: 675–684.
http://dx.doi.org/10.1016/0192-0561(90)90105-V
Tafalla, C., Novoa, B (2000). Requirements for nitric oxide production by turbot (Scophthalmus maximus) head kidney macrophages. Dev. Comp. Immunol., 24: 623–631.
http://dx.doi.org/10.1016/S0145-305X(99)00087-7
Tayade C, Koti M and Mishra SC (2006). L–arginine stimulates intestinal intraepithelial lymphocyte functions and immune response in chickens immunized with intermediate plus strain of infectious bursal disease vaccine. Vaccine 24: 5473–5480.
http://dx.doi.org/10.1016/j.vaccine.2006.03.086
http://dx.doi.org/10.1016/j.vaccine.2005.08.059
Tominac VP, Krpan VZ, Grba S, Srecec S, Krbavcic IP and Vidovic L (2010). Biological effects of yeast beta glucans. Agri. Con. Sci. 75: 149–158.
Verlhac V, Obach A, Gabaudan J, Schuep W and Hole R (1998). Immunomodulation of Dietary vitamin C and glucan in rainbow trout. Fish Shellfish Immunol. 8: 409–424.
http://dx.doi.org/10.1006/fsim.1998.0148
Vetvicka V, Vashishta A, Saraswat–ohri S and Vetvickova J (2008). Immunological effects of yeast– and mushroom–derived beta–glucans. J. Med. food 11: 615–622.
http://dx.doi.org/10.1089/jmf.2007.0588
PMid:19053851
Weickert MO and Pfeiffer AF (2008). Metabolic effects of dietary fiber consumption and prevention of diabetes. J. Nutr. 138: 439–442.
PMid:18287346
Xing Z and Schat KA (2000). Expression of cytokine genes in Marek's disease virus –infected chicken and chicken embryo fibroblast cultures. Immunol. 100:70–76.
http://dx.doi.org/10.1046/j.1365-2567.2000.00008.x
PMCid:PMC2326989
Yoshida T., Sakai M, Kitao T, Khill SM, Araki S, Saito R, Ineno T and Inglis,V (1993). Immunomodulatory effects of the fermented products of chicken egg, EF203, on rainbow trout, Onchorhynchus mykiss. Aquacult. 109:207–214.
http://dx.doi.org/10.1016/0044-8486(93)90163-S

