

Seasonal Variation in the Trophic Structure of the Fish Community in Dongshan Bay and Adjacent Waters





Seasonal Variation in the Trophic Structure of the Fish Community in Dongshan Bay and Adjacent Waters
Ying Guan1,2, Puqing Song2, Xing Miao2, Hai Li2, Rui Wang2, Liangming Wang3, Yuan Li2* and Longshan Lin2*
1College of Environmental Science and Engineering, Ocean University of China, Qingdao, P.R. China, 266100
2Third Institute of Oceanography, Ministry of Natural Resources, Xiamen, P.R. China, 361005
3South China Sea Fishers Research Institute, Chinese Academy of Fisheries Sciences, Guangzhou, P.R. China, 510300
ABSTRACT
Based on bottom trawl survey data collected in November 2009 (autumn), February 2010 (winter), May 2010 (spring), and August 2010 (summer), the seasonal variation in the trophic structure of the fish community in Dongshan Bay and adjacent waters was investigated. A total of 114 species of fish were caught in surveys throughout the year. According to their feeding habits, the fish species in the surveyed area were classified into six functional groups, including planktivores (FG1), planktivores/benthivores (FG2), benthivores (FG3), benthivores/piscivores (FG4), piscivores (FG5), and omnivores (FG6). FG3 was the most diverse group, with 43 species, followed by FG4, with 29 species. The biomass of FG3 was the highest among all functional groups throughout the year and in each individual season. FG1 and FG5 had higher biomass proportions in spring than in the other seasons, accounting for 8.02% and 16.72%, respectively. The functional group diversity (HFD) and species diversity (Hs) showed similar variations, reaching their lowest values in winter and highest values in summer. In 2009-2010, the mean trophic position of the fish community in Dongshan Bay and adjacent waters was 3.64. This value was 3.74 in winter (highest), 3.49 in summer (lowest), 3.71 in spring, and 3.63 in autumn. The relationship between the trophic position and the mean body mass indicated that the seasonal variation in the mean body mass of high-trophic-level fish species was the highest in winter, followed by autumn, spring, and summer. In general, with seasonal and temporal changes, large-bodied, high-trophic-level fishes were gradually replaced by small-bodied, low-trophic-level fishes in the fish community of Dongshan Bay and adjacent waters in 2009-2010, and the trophic structure tended to become simplified.
Article Information
Received 10 January 2022
Revised 18 February 2022
Accepted 15 March 2022
Available online 15 June 2022
(early access)
Published 12 June 2023
Authors’ Contribution
PQS, YL and LSL conceived and designed. XM and LMW conducted the survey. RW collected background information. YG wrote and edited the article. All authors read and approved the final manuscript.
Key words
Dongshan bay, Nutrition structure, Functional guilds, Mean trophic position, Seasonal variation
DOI: https://dx.doi.org/10.17582/journal.pjz/20220110010121
* Corresponding author: [email protected]; [email protected]
0030-9923/2023/0004-1691 $ 9.00/0
Copyright 2023 by the authors. Licensee Zoological Society of Pakistan.
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
INTRODUCTION
Fish occupy a broad position in the aquatic food web of the ocean. Fish community structures can exhibit apparent changes within and across seasons (Xu and Shen, 2011). The trophic position (TL) can be used as an indicator to measure the system stability and resource development level. As the TL concept has continued to develop, research on TLs and the trophic structure in recent years has gradually gained importance in the fields of contemporary fishery ecology and marine ecodynamics (Ji et al., 2010). In ecological research, the assignment of functional groups simplifies the interpretation of interspecific relationships and weakens the effects of individual species within communities, which can make research on ecosystems more concise and enhance the stability and predictability of research on communities (Steneck and Watling, 1982).
Bay ecosystems are a special natural system composed of semi-enclosed water bodies and associated biological communities, which are surrounded by land along the coast (State, 2005). Bays have complex environments and diverse habitats and are most affected by human activities given their unique geographic locations. In recent years, studies of nekton in Dongshan Bay and adjacent waters in China have mainly focused on the abundance and distribution characteristics of fish eggs, larvae, and juveniles, the diversity of nekton, the community structure, and the fishery resource carrying capacity (Zhang et al., 2013a; Chen, 2016; Cai et al., 2021). Yan et al. (1990) investigated the fish composition and its seasonal variation in Dongshan Bay. Lin et al. (2012 a, b), Zhang et al. (2013a, b), Wu (2016), and Ye et al. (2018) recently analyzed the fish taxonomic diversity and community structure in Dongshan Bay and its adjacent waters. Ye et al. (2018) and Zhang et al. (2013a, b, c) evaluated and analyzed indicators of the fishery resource carrying capacity and aquaculture carrying capacity of Dongshan Bay. However, studies on the feeding habits and TLs of fish in this region is limited to studies by Zhang and others (Zhang et al., 1993, 1994; Zhang, 1996) on the fish food web in Dongshan Bay and the feeding habits of commercial fish species, including Sebastiscus marmoratus and Johnius grypotus.
In this study, vessel survey data from four seasons, gathered in November 2009, February 2010, May 2010, and August 2010, were used to conduct species identification of fish samples. By combining our data with the relevant literature, we analyzed the seasonal variation in the trophic structure of the fish community in Dongshan Bay. To the best of our knowledge, this is the first study to report on the assignment of functional groups to fishes in Dongshan Bay and the seasonal variation in the TLs of its fishes. This study aimed to generate fundamental information for future research on the food web and trophic dynamics of Dongshan Bay to serve as a reference for the development and maintenance of fishery resources of the bay.
MATERIALS AND METHODS
Survey area and stations
Dongshan Bay is located on the west side of the Taiwan Strait and in the southernmost part of Fujian Province. It is a semienclosed inner bay surrounded by the coasts of Zhangpu County, Yunxiao County, and Dongshan County in Zhangzhou City. Dongshan Bay is the largest bay in southern Fujian, and given its geographical location, complex topography, landforms, and unique ecological environment, it provides an optimal environment for fishes to inhabit, reproduce, and grow. Many fish species congregate in Dongshan Bay in different seasons to reproduce, feed, and grow in addition, Dongshan Bay is one of the three main bases of fishery production in Fujian Province (Zhou, 2002). With the rapid industrial development and urbanization of the areas surrounding Dongshan Bay, the deterioration of the marine environment has intensified, and the self-regulating ability of the Dongshan Bay ecosystem has declined substantially, thereby greatly threatening its marine ecosystem (Yan et al., 2019).
Study data were collected from four vessel surveys, which were conducted in November 2009 (autumn), February 2010 (winter), May 2010 (spring), and August 2010 (summer) in Dongshan Bay and adjacent waters. A total of 16 stations were set up in the vessel surveys and distributed as shown in Figure 1. Among these stations, five were in the bay (stations 1 to 5), and 11 were outside of the bay (stations 6 to 16).
Data collection
The survey vessel was a winged single-net bottom trawler assembled with a fishery cod-end net with a swept width of 14 m and a mesh size of 20 mm.
Based on the sediment quality in the surveyed waters and the surrounding environment, the trawl time for each station was approximately 30 min at a trawl speed of 2.5-3.2 kn. The fish were sampled, weighed, enumerated, and measured anatomically in accordance with Specifications for Oceanographic Survey-Part 6: Marine Biological Survey (GB/T12763.6-2007) (General et al., 2007). Fish species caught in Dongshan Bay and adjacent waters were identified with reference to Fishes of Fujian Province (Zhu et al., 1984).
Statistical analysis
The diversity index and functional group index were both calculated using the following equation (Shannon et al., 1949):
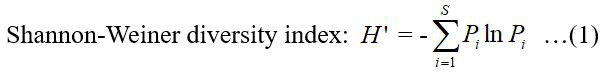
Where, Pi is the proportion of biomass of fish species i to the total fish biomass. When calculating the functional group diversity, Pi is the proportion of biomass of functional group i to the total fish biomass.
The mean TL (MTL) of the fish community was analyzed using the following equation (Pauly et al., 1998):
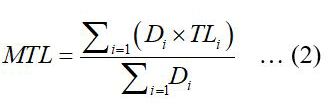
Where Di is the relative biomass of fish species i (kg/h), and TLi is the TL of fish species i.
According to the current standard convention, the TL of primary producers was set to 1 (Yang, 1982). TLs range from 2.4-2.9 for omnivorous fishes, 3.0-3.8 for low-level carnivorous fishes, 3.9-4.4 for middle level carnivorous fishes, and 4.5 and up for high-level carnivorous fishes (Zhang et al., 1981). To classify the TLs of the fishes in Dongshan Bay, we mainly referred to studies on fish TLs in the coastal waters of Fujian Province (Lu and Huang, 2004) and fish food webs in waters east of Xiamen (Zhang and Tang, 2004) and in Dongshan Bay (Zhang et al., 1994). Next, the TL classification of fishes was adjusted according to standard conventions. For fish species not found in the aforementioned literature, TLs were assigned by referencing FishBase.
The diversity index, functional group index, TL, and mean body mass were calculated in Excel 2016. The correlation between the TL and the mean body mass was also calculated. Using R 3.6.2, seasonal and spatial differences in the diversity index and the functional group diversity index were tested using one-way analysis of variance (ANOVA), followed by Duncan’s multiple range test, and the results were used to graphically present seasonal and spatial differences.
RESULTS
Species composition of each functional group
From the four vessel surveys of nekton in Dongshan Bay and adjacent waters, a total of 114 species of fish, belonging to 15 orders, 57 families, and 81 genera, were caught. Based on the feeding habits of the fishes, six functional groups were identified: planktivores (FG1), planktivores/benthivores (FG2), benthivores (FG3), benthivores/piscivores (FG4), piscivores (FG5), and omnivores (FG6). FG3 has the largest number of species, with 43 (Fig. 2A), including Narke japonica, Platyrhina limboonkengi, and Evynnis cardinalis, accounting for 37.72% of all species. FG4 was the second-largest group, with 29 species, including Johnius grypotus and Nemipterus virgatus, accounting for 25.44%. FG6 was the smallest group, with seven species, including Trachurus japonicus and Hapalogenys nitens, accounting for only 6.14%.
The species composition of each functional group in different seasons was consistent with the annual composition (Fig. 2B). However, the relative proportion of FG3 and FG6 increased considerably in summer, whereas the relative proportion of FG4 showed a decreasing trend from autumn to summer.
Biomass composition of functional groups
In terms of biomass (Fig. 3A), FG3 was the dominant group (accounting for 63.66% of the total), followed by FG4, FG5, FG6, FG1, and FG2. Seasonal variations in the biomass composition of the functional groups were apparent (Fig. 3B). FG3, which mostly included N. japonica, P. limboonkengi, and E. cardinalis, was still the dominant group in each season. In particular, the biomass of FG3 was substantially greater in autumn than in the other seasons due to the presence of more N. japonica and P. limboonkengi. Owing to the greater occurrence of Thryssa mystax and Saurida elongata, the relative biomass proportion of FG1 and FG5 was greater in spring than in other seasons. FG6 had a fair biomass proportion in summer but a very low proportion in the other seasons.
Functional group diversity
The seasonal variations in the functional group diversity (HFD) and the species diversity (HS) were similar, and both were the highest in summer and the lowest in winter, exhibiting an increasing trend overall (Fig. 4). One-way ANOVA with multiple comparisons showed that HFD did not have significant differences among seasons (P > 0.05), while HS did not differ significantly between spring and summer or between autumn and winter (P > 0.05), while other pairwise comparisons between seasons revealed significant differences (P < 0.05).
Seasonal variation in MTL
With reference to the relevant literature (Zhang et al., 1981, 1994; Lu and Huang, 2004; Huang et al., 2008; Zhang and Tang, 2004; Wei and Jiang, 1992) and FishBase, we determined the TLs of 114 fish species in Dongshan Bay (Table I, the TL of main species). The MTL in Dongshan Bay and adjacent waters was calculated for all seasons (Table II). The MTL of the fish community in Dongshan Bay and adjacent waters was 3.64, which is an intermediate level, with limited seasonal variations. The MTL of the fish community in summer (3.49) was slightly lower than that in autumn (3.63), winter (3.74), and spring (3.71). The MTL of the fish community in Dongshan Bay exhibited an increasing trend from autumn to winter and an overall decreasing trend from winter to summer, with a relatively large reduction in summer.
Seasonal variation in TL and mean body mass
The TL–mean body mass relationship curve reflected variations in the mean body mass of the fish in each TL and the fish community structure (Wang et al., 2012). Using TL ranges of <3.5, 3.5-3.9, and >3.9, the fish samples collected from Dongshan Bay in all four seasons in 2009–2010 were classified into three categories. Figure 5 in autumn, the mean body mass of fishes with a TL below 3.5 was 13.87 g, that of fishes with a TL between 3.5 and 3.9 was 66.46 g, and that of fishes with a TL greater than 3.9 was 458.24 g; in winter, the mean body mass of the fish samples in these three TL categories was 57.19 g, 130.81 g, and 273.05 g, respectively; in spring, the mean body mass of the fishes in these three TL categories was 28.21 g, 89.15 g, and 202.39 g, respectively; and in summer, the mean body mass of the fishes in these three TL categories was 24.37 g, 104.57 g, and 115.69 g, respectively. The mean body mass of the fishes with high TLs (i.e., greater than 3.9) showed an apparent decreasing trend across seasons, whereas that of the fishes with TLs below 3.5 increased significantly in winter.
Table I. Trophic position and functional guild of fishes in Dongshan Bay.
|
Functional guilds/ Species |
Trophic position |
References |
|
FG1: Planktivores |
||
|
Sardinella aurita |
3.10 |
(Zhang and Tang, 2004) |
|
Bleekeria anguilliviridis |
3.20 |
* |
|
Anchoviella commersonii |
3.20 |
(Huang and Zhang, 2008) |
|
Thryssa mystax |
3.40 |
(Huang and Zhang, 2008) |
|
FG2: Planktivores/Benthivores |
||
|
Setipinna tenuifilis |
3.10 |
(Zhang and Tang, 2004) |
|
Leiognathus ruconius |
3.10 |
(Huang and Zhang, 2008) |
|
Leiognathus bindus |
3.30 |
(Zhang and Tang, 2004) |
|
Psenopsis anomala |
3.50 |
(Zhang and Tang, 2004) |
|
Apogon lineatus |
3.80 |
* |
|
Decapterus maruadsi |
3.80 |
(Huang and Zhang, 2008) |
|
FG3: Benthivores |
||
|
Siganus punctatus |
2.40 |
* |
|
Evynnis cardinalis |
3.10 |
(Huang and Zhang, 2008) |
|
Leiognathus rivulatus |
3.30 |
(Zhang and Tang, 2004) |
|
Takifugu bimaculatus |
3.30 |
* |
|
Thrissa dussumieri |
3.40 |
(Zhang and Tang, 2004) |
|
Takifugu alboplumbeus |
3.40 |
(Zhang and Tang, 2004) |
|
Narke japonica |
3.50 |
* |
|
Platyrhina limboonkengi |
3.50 |
* |
|
Acanthopagrus berda |
3.50 |
* |
|
Upeneus bensasi |
3.50 |
(Zhang and Tang, 2004) |
|
Terapon theraps |
3.60 |
(Huang and Zhang, 2008) |
|
Argyrosomus argentatus |
3.60 |
(Zhang and Tang, 2004) |
|
Sillago sihama |
3.60 |
(Huang and Zhang, 2008) |
|
Argyrosomus argentatus |
3.60 |
(Zhang and Tang, 2004) |
|
Priacanthus macracanthus |
3.60 |
(Zhang and Tang, 2004) |
|
Acanthopagrus latus |
3.60 |
(Zhang and Tang, 2004) |
|
Raja hollandi |
3.70 |
(Zhang et al., 1994) |
|
Polydactylus sextarius |
3.80 |
(Huang and Zhang, 2008) |
|
Platyrhina sinensis |
4.00 |
(Zhang et al., 1994) |
|
Dasyatis zugei |
4.00 |
(Zhang and Tang, 2004) |
|
Dasyatis akajei |
4.00 |
(Zhang and Tang, 2004) |
|
Ophichthus apicalis |
4.10 |
(Zhang and Tang, 2004) |
|
Raja kenojei |
4.20 |
(Zhang and Tang, 2004) |
|
Cynoglossus joyneri |
4.50 |
* |
|
Functional guilds/ Species |
Trophic position |
References |
|
FG4: Benthivores/Piscivores |
||
|
Takifugu oblongus |
3.30 |
(Huang and Zhang, 2008) |
|
Chelidonichthys kumu |
3.30 |
(Huang and Zhang, 2008) |
|
Gastrophysus spadiceus |
3.30 |
(Huang and Zhang, 2008) |
|
Harpadon nehereus |
3.50 |
(Huang and Zhang, 2008) |
|
Johnius grypotus |
3.50 |
(Huang and Zhang, 2008) |
|
Lepidotrigla kishinouyi |
3.50 |
* |
|
Arius sinensis |
3.50 |
(Huang and Zhang, 2008) |
|
Sebastiscus marmoratus |
3.60 |
(Zhang and Tang, 2004) |
|
Apogon quadrifasciatus |
3.60 |
(Huang and Zhang, 2008) |
|
Gymnura japonica |
3.90 |
* |
|
Lagocephalus inermis |
3.90 |
* |
|
Chiloscyllium plagiosum |
4.00 |
(Zhang and Tang, 2004) |
|
Gymnura bimaculata |
4.00 |
* |
|
Lateolabrax japonicus |
4.40 |
(Huang and Zhang, 2008) |
|
Nemipterus virgatus |
4.60 |
(Huang and Zhang, 2008) |
|
FG5: Piscivores |
||
|
Pisoodonophis cancrivorus |
3.80 |
(Zhang and Tang, 2004) |
|
Epinephelus akaara |
4.00 |
* |
|
Saurida elongata |
4.20 |
(Huang and Zhang, 2008) |
|
Dasyatis bennetti |
4.50 |
* |
|
Trichiurus japonicus |
4.60 |
(Huang and Zhang, 2008) |
|
FG6: Omnivores |
||
|
Leiognathus lineolatus |
3.10 |
(Zhang and Tang, 2004) |
|
Scomber japonicus |
3.50 |
* |
|
Trachurus japonicus |
3.70 |
(Zhang and Tang, 2004) |
Note: (Zhang et al., 1994; Yang, 1982; Zhang et al., 1981; Lu and Huang, 2004; Huang et al., 2008; Zhang and Tang, 2004) meant data gaining from the relative articles, and “*” meant data gaining from Fishbase.
Table II. Mean trophic position of fish species in Dongshan Bay from 2009 to 2010.
|
Autumn |
Winter |
Spring |
Summer |
Total |
|
|
Mean trophic position |
3.63 |
3.74 |
3.71 |
3.49 |
3.64 |
DISCUSSION
Functional groups
Stable ecosystems should have a relatively large number of species, a complex food web structure, functional groups that are composed of many species that can substitute for each other, and diverse pathways of material and energy flows. Although the FG3 had a high species composition and biomass in this study, however, compared with the findings of Zhang et al. (1994) in the 1990s, the proportion of FG3 in this study decreased from 41.67% to 37.72%, which could be due to the presence of demersal cartilaginous fishes, such as N. japonica, P. limboonkengi, and Platyrhina sinensis. These fishes have weak swimming abilities and mainly feed on benthic invertebrates, such as Macrura Reptantia, mysids, cumaceans, and amphipods (Yuan et al., 2017). The proportion of FG4 increased considerably from 15.48% to 25.44%, which could be associated with the presence of large demersal cartilaginous fishes, such as Gymnura japonica and Gymnura bimaculata, or with the transition in feeding habits from benthivory to benthivory/piscivory, such as in Sebastiscus marmoratus.
The species composition and biomass of FG6 increased significantly in summer compared to the other seasons. Some omnivorous fish species, including Scomber japonicus and Aluterus monoceros, are only present in summer. S. japonicus is a mesopelagic migratory fish that makes vertical diel migrations. In winter, S. japonicus populations inhabit deeper waters, and in summer, when the water temperature has increased, S. japonicus populations migrate to coastal areas to spawn. In spring and summer, FG6 was dominated by T. japonicus, which is an important species in summer (index of relative importance=702) (Zhang et al., 2013a). And this is probably because the main prey items of T. japonicus zooplankton proliferate in spring and summer. T. japonicus is a warm-water migratory fish, the larvae of which mainly feed on small-bodied zooplankton, including copepods, cladocerans, krill, and mysids. From the juvenile to the adult stage, T. japonicus feeds on zooplankton, such as krill, mysids, crustacean larvae, and sardine larvae, as well as small-bodied fishes and cephalopods (Cheng and Zhou, 1997).
The functional group diversity and the species diversity both showed their lowest values in autumn, when the stability of the community structure became weak. The functional group diversity was lower than the species diversity, and the seasonal variation of the former was limited. At most stations, both the functional group diversity and the species diversity were relatively low. In this study, the six functional groups had considerable differences in species richness and biomass. The relatively wide seasonal variations in the functional groups (e.g., FG6 and the benthivores in FG3) and their relatively low diversity indicated that the community trophic structure was unstable. In recent years, rapid industrial and aquacultural development in Dongshan Bay has caused serious pollution in the aquatic environment, leading to changes in the community structure, which then lead to variations in the trophic structure (Chen et al., 1994).
Seasonal variation in MTL
The accumulation and transfer of energy and biomass between TLs are key steps of maintaining ecosystem stability. The quantitative relationships between energy and matter between TLs are the material basis of an ecosystem (Elton, 1930). The MTL of the marine catch (also known as the marine trophic index) is one of the eight diversity indices used to directly evaluate biological diversity (Pauly and Watson, 2005). In this study, based on catch data obtained from surveys in Dongshan Bay, the trophic structure of the fish community and its variations in 2009-2010 were analyzed. We found that the annual MTL was 3.64, between 3.49 (in summer) and 3.74 (in winter). In winter, the proportions of P. limboonkengi, with a medium to high TL (TL=3.50), in FG3 and Hemitrygon bennettii (TL=4.50) in FG5 were relatively high, at 22.44% and 14.76%, respectively. In summer, the MTL of the fish community decreased, mainly because E. cardinalis, with a medium to low TL (TL=3.10), accounted for the highest proportion of FG3, at 24.41%, and Siganus oramin, with a low TL (TL=2.40), also accounted for a relatively high proportion of 5.35%. E. cardinalis is a warm-water, small-bodied demersal migratory fish, with a spawning period from March to April. Therefore, in summer, the abundance of this small fish increases substantially, leading to a decrease in the fish MTL in summer. Under the influence of overfishing, the abundance of large-bodied, high-trophic-level predators with greater economic values continues to decline, resulting in a transition in the catch composition; i.e., more small-bodied, lower trophic level fishes with lower economic values are caught. Such changes in the fish community structure ultimately lead to changes in the fish MTL (Pauly et al., 1998).
Seasonal variation in the TL mean body mass curve
According to Yuan et al. (2017), a greater slope of the TL mean body mass curve indicates a higher mean body mass of fishes with high TLs and vice versa. The TL mean body mass relationship curves for all seasons showed that the mean body mass of the high trophic level fishes was highest in winter, followed by autumn, spring, and summer. The mean body mass of the fishes with a TL greater than 3.9 was 458.24 g in the autumn of 2009, which was significantly higher than that (273.05 g) in winter. The main reason was that the catch in the autumn included large bodied, high trophic level Trichiurus japonicus (TL=4.60), Cynoglossus joyneri (TL=4.50), and Arius thalassinus (TL=4.50), which had mean body masses of 908.00 g, 598.50 g, and 331.33 g, respectively. In this study, the mean body mass of the high trophic level fishes in spring and summer was lower than that in autumn and winter. Spring and summer are the reproductive migration and spawning periods of fishes, so the abundance of small, high trophic level fishes, such as T. japonicus and S. elongata (TL=4.20), increases substantially. Water temperatures are low in autumn and winter, so the metabolism of fish is significantly slower, and their swimming ability is also weak. Therefore, the high-trophic-level adult fishes that are caught often have a relatively large body size. The dominant species also exhibited seasonal changes. From autumn and winter to spring and summer, the dominant species shifted from long life cycles to short life cycles, from high TLs to low TLs, from high quality to low quality, and from large medium body sizes to medium small body sizes (Pauly et al., 1998).
Dongshan Bay is a subtropical sea area that includes many islands, and it originally had rich biological resources. However, fishery resources in this area have experienced gradual declines in recent years due to marine development and utilization and rapid growth of the economy and the population, particularly the rise of fishing and aquaculture industries. In this study, only the trophic structure of the fish community and its seasonal variation in Dongshan Bay and adjacent waters were investigated within a relatively short timeframe. Long-term monitoring of Dongshan Bay and its adjacent waters is needed for more in-depth studies of the variation patterns in the fish food web and TLs in this area. To conclude, this study showed that under the effects of environmental pollution caused by overfishing and marine development and utilization, medium to large high trophic level fishes tend to be gradually replaced by small low-trophic-level fishes in the Dongshan Bay fish community and this trend need to be verified by longer time scale studies. With seasonal and temporal changes, the community MTL decreased, and the trophic structure of the fish community became simplified.
CONCLUSION
In this study, 114 fish species were classified into six functional groups, and the biomass of benthivores (FG3) was the highest among all functional groups throughout the year and in each individual season. The low values of functional group diversity (HFD) and species diversity (HS) may indicate the instability of the trophic structure in the study area. The annual MTL ranged from 3.49 (in summer) to 3.74 (in winter), which may be caused by the seasonal varies in the biomass of some species at low trophic levels, such as E. cardinalis etc. The relationship between the trophic position and the mean body mass indicated that the seasonal variation in the mean body mass of high-trophic-level fish species was the highest in winter, followed by autumn, spring, and summer. In general, the community structure of nekton in Dongshan Bay is in dynamic change. On the one hand, the seasonal variation of the abundance of some small fishes affects the trophic structure of the sea area, on the other hand, under the long-term influence of human factors such as fishing and aquaculture, the biological composition of Dongshan Bay changes significantly, which leads to the interannual change of trophic structure.
ACKNOWLEDGEMENTS
The present study could not have been performed without assistance from Miss Yuanyuan Li during the writing of the manuscript. We also thank the anonymous reviewers for their helpful comments. This work was supported by the National Key Research and Development Program of China (2018YFC1406302), the Scientific Research Foundation of TIO, MNR (2019017, 2019018). The authors declare no conflicts of interest including the implementation of research experiments and writing this manuscript.
Statement of conflict of interest
The authors have declared no conflict of interest.
REFERENCES
Cai, J.D., Xu, C.Y., Ma, C., Ye, S.Z., Zhuang, X.D., Liu, Y., Xie, S.Q., and Shen, C.C., 2021. Population aggregation characteristics of the main dominant species of nekton in Minnan fishing ground. Acta Ecol. Sin., 41: 4818-4826. https://doi.org/10.5846/stxb202001070052
Chen, C.Q., Ruan, J.S., Li, X.Z., Hong, Y.H. and Hong, G.Y., 1994. Investigation report on fishery environment monitoring in Dongshan Bay. J. Fish. Res., 3: 56-65.
Chen, J., 2016. Distribution pattern of taxonomic diversity of fish community in four typical mangrove areas, China. Xiamen University, Xiamen.
Cheng, Q.T. and Zhou, C., 1997. The fishes of Shandong Province. Science and Technology Press, Shandong, China.
Elton, C.S., 1930. Animal ecology and evolution. Nature, 128: 2223-2224.
General administration of quality supervision, inspection and quarantine of the people’s republic of China, standardization administration of the People’s Republic of China, 2007. Specifications for Oceanographic Survey- Part 6: Marine Biological Survey (GB/T12763.6-2007). Standards Press of China, Beijing, China.
Huang, L.M., Zhang, Y.Z., Pan, J.J., Cui, J.J. and Wu, Y., 2008. Food web of fish in Xiamen eastern waters. J. appl. Oceanogr., 27: 64-73.
Huang, Y., 1999. Species composition and seasoned variations of fishes in Dongshan Bay. J. Fish. Res., 2: 55-61.
Ji, W.W., Li, S.F. and Chen, X.Z., 2010. Application of fish trophic position in marine eco-system. J. Fish. Sci. China, 17: 878-887.
Lin, L.S., Wang, Y.P., Li, Y., Zhang, J. and Gao, T.X., 2012. Taxonomic diversity of fish species in Dongshan Bay, China and adjacent areas. J. Fish. Sci. China, 19: 1060-1067. https://doi.org/10.3724/SP.J.1118.2012.01060
Lin, L.S., Zhao, G.G., Li, Y., Gao, T.X. and Zhang, J., 2012. Diversity of nekton in Dongshan Bay and adjacent areas. Acta Hydrobiol. Sin., 36: 1119-1127. https://doi.org/10.3724/SP.J.1035.2012.01119
Lu, Z.B. and Huang, M.Z., 2004. Studies on frophic level and organic carbon contents of main economic nekton in Fujian coastal waters. J. appl. Oceanogr., 23: 153-158.
Pauly, D. and Watson, R., 2005. Background and interpretation of the marine trophic index as a measure of biodiversity. Phil. Trans. R. Soc. B., 360: 415-423. https://doi.org/10.1098/rstb.2004.1597
Pauly, D., Christensen, V. and Dalsgaard, J., 1998. Fishing down marine food webs. Science, pp. 279. https://doi.org/10.1126/science.279.5352.860
Shannon, C.E., Weaver, W. and Wiener, N., 1949. The mathematical theory of communication. Bell. Labs. Tech. J., 3: 31-32. https://doi.org/10.1063/1.3067010
State Oceanic Administration, 2005. The guidance for the assessment of coastal marine ecosystem health. Standards Press of China, Beijing, China.
Steneck, R.S. and Watling, L., 1982. Feeding capabilities and limitation of herbivorous molluscs: A functional group approach. Mar. Biol., 68: 299-319. https://doi.org/10.1007/BF00409596
Wang, Y.B., Zheng, J., Zheng, X.Z. and Qin, T., 2012. Variation analysis of single otter trawl fish community structure outside forbidden fishing line of Zhoushan fishing ground. South. China Fish. Sci., 8: 8-15.
Wei, S. and Jiang, W.M., 1992. Study on food web of fish in the Yellow Sea. Oceanol. Limnol. Sin., 2: 182-192.
Wu, H.R., 2016. Survey on status of fishery resources of Dongshan Bay in Fujian. J. Fish. Res., 38: 32-39.
Xu, Z.L. and Shen, A.L., 2011. Spatial and temporal distribution on fish diversity in the Xinghua Bay. J. Fish. Sci. China, 18: 416-426. https://doi.org/10.3724/SP.J.1118.2011.00416
Yan, L., Fang, J., Wang, J.K. and Wu, Y.J., 2019. The scope of spatial planning of dongshan bay coastal wetland in Fujian Province. Wetland Sci. Manage., 15: 55-58.
Yan, Y.M., Dai, Q.S. and Lu, Z.B., 1990. Study on the structure and rational utilization of fish resources in the waters around Dongshan Island. J. Fish. Res., 3: 32-39.
Yang, J., 1982. A tentative analysis of the trophic positions of North Sea fish. Mar. Ecol. Prog., 7: 247-252. https://doi.org/10.3354/meps007247
Ye, S.Z., Luo, D.L. and Cai, J.D., 2018. Characteristics of community structure and biomass distribution of fishery resources in Dongshan Bay. J. Fish. Res., 40: 358-365.
Yuan, H.R., Chen, P.M., Qin, C.X., Lin, X.G., Zhou, Y.B. and Feng, X., 2017. Seasonal variation of fish community structure in Zhelin Bay, the South China Sea. South China Fish. Sci., 13: 26-35.
Zhang, B. and Tang, Q.S., 2004. Study on trophic position of important resources species at high trophic position in the Bohai Sea, Yellow Sea and East China Sea. Adv. Mar. Sci., 4: 392-404.
Zhang, J., Song, P.Q., Chen, Y.J., Zhong, Z.H. and Lin, L.S., 2013a. Species composition and abundance temporal-spatial distribution of ichthyoplankton in Dongshan Bay of Fujian. J. Fish. Res., 35: 1-7.
Zhang, J., Xu, S.J., Li, Y., Gao, T.X., Song, P.Q. and Lin, L.S., 2013b. Studies on fish community structure in Dongshan Bay and its adjacent areas. Period. Ocean. Univ. China., 43: 51-57.
Zhang, J., Chen, Y.J., Song, P.Q., Zhong, Z.H. and Lin, L.S., 2013c. Nekton composition and diversity of Dongshan Bay, Fujian. Mar. Fish., 35: 15-23.
Zhang, Q.Y., Lin, Q.M., Lin, Y.T. and Zhang, Y.P., 1981. Food web of fishes in Minnan-Taiwanchientan fishing ground. Acta Oceanol. Sin., 3: 275-290.
Zhang, Y.Z., 1996. Study on feeding habits of Johnius belengerii in Dongshan Bay. J. Jimei. Univ., 18: 25-32.
Zhang, Y.Z., Li, F.Z. and Guo, C.C., 1993. Study on feeding habits of Sebastiscus marmoratus in Dongshan Bay. J. appl. Oceanogr., 12: 233-241.
Zhang, Y.Z., Li, F.Z., Liu, X.Y., Lan, X.W. and Chen, J.K., 1994. Food web of fishes in Dongshan Bay, Fujian. J. appl. Oceanogr., 1: 52-61.
Zheng, H.D., 2016. Abundance distribution of fish eggs and larvae and its relation with environmental factors in spring and summer in Dongshan Bay, Fujian. J. appl. Oceanogr., 35: 87-94.
Zheng, W.B. and Qiu, S.Y., 1989. Feeding habits of cartilaginous fishes. J. Fish. Res., 4: 56-61.
Zhou, Q.L., 2002. The biodiversity of four typical zones in the south-east ocean of China: The 5th national symposium on biodiversity conservation and sustainable utilization. Meteorological Press, Beijing, China.
Zhu, Y.D., Wu, H.L. and Jin, X.B., 1984. The fishes of Fujian Province. Fujian Science And Technology Press, Fuzhou, China.
To share on other social networks, click on any share button. What are these?


