
The Journal of Advances in Parasitology


Short Communication
The Journal of Advances in Parasitology 1 (3): 41 – 43Polymorphism at the Merozoite Surface Protein–3α Locus of Plasmodium vivax in Sri Lanka
Yoshitha Wahalathanthri1, Gawrie Galappaththy2, Deepika Fernando3, Rabindra Abeyasinghe2, Neil Fernandupulle1, Dammika Senevirathna1*
- Genetech Research Institute, No.54, Kithulwatta Road, Colombo–08, Sri Lanka
- Anti Malaria Campaign, No: 555/5, Public Health Building, Narahenpita, Sri Lanka
- Faculty of Medicine, University of Colombo, Sri Lanka
*Corresponding author:dbsenevi22@gmail.com
ARTICLE CITATION:
Wahalathanthri Y, Galappaththy G, Fernando D, Abeyasinghe R, Neil Fernandupulle N, Senevirathna D (2014). Polymorphism at the merozoite surface protein–3α locus of plasmodium vivax in Sri Lanka. J. Adv. Parasitol. 1 (3): 41 – 43.
Received: 2014–07–03, Revised: 2014–08–14, Accepted: 2014–08–15
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/journal.jap/2014/1.3.41.43
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
The knowledge on the population structure of Plasmodium vivax isolates has contributed greatly to understanding the dynamics of disease transmission. Here we assess the potential of parasite transmission in Sri Lanka using genetic diversity of a highly polymorphic marker MSP3α. A total of 26 cases presenting with microscopically confirmed asexual P.vivax mono–infections during the period January to December 2011 were analysed. Six distinct allele variants were identified using PCR/RFLP analysis of PVMSP3α with HhaI. The present data indicates that a comparatively low level of diversity among P.vivax populations exists in Sri Lanka and this could be a contributing factor towards the success of eliminating of malaria from the country by 2014.
Despite enormous control efforts over many decades malaria still remains one of the most important communicable diseases in the world (WHO, 2013). In 2009, the Ministry of Health’s Anti Malaria Campaign, Sri Lanka entered the malaria pre–elimination phase as the number of indigenous malaria cases reported was less than 200 per year (AMC, 2011). With no indigenous malaria cases being reported since October 2012, Sri Lanka has targeted to achieve its goal of malaria elimination by the end of 2014. The genetic diversity and population structure of P. vivax have a significant impact on malaria transmission, spread of drug–resistance and the acquisition of protective immunity against malaria (Hajime et al., 2011). Therefore, this study was designed to find out the genetic diversity and population structure of P. vivax in a cohort of isolates collected from patients presenting with malaria during the period January to December 2011 in order to determine the strength towards eliminating P.vivax from the country by considering the changing pattern of genetic diversity. A polymorphic genetic marker, the merozoite surface protein–3α (MSP–3α) was selected to investigate the population diversity of P. vivax using Polymerase Chain Reaction–Restriction Fragment Length Polymorphism (PCR–RFLP) analysis of the highly polymorphic PVMSP3α gene. Diversity studies with isolates from around the world continue to show extensive polymorphism in PVMSP3α and the encoded protein, and the PVMSP3α alleles are therefore highly regarded as a genetic tool to distinguish different parasite isolates and study population dynamics (Schousboe et al., 2011).
Of the 119 P.vivax cases reported in 2011, 26 (21.84 % of the total infected cases) cases presenting with microscopically confirmed asexual P.vivax mono–infection were used to assess the genetic diversity. None of the patients gave a history of travel abroad. Finger prick blood was collected under aseptic conditions to Whatman Filter Paper 3 prior to starting anti malaria treatment. Ethical clearance was obtained from the Ethics Review Committee, Sri Lanka Medical Association.
The parasite DNA was extracted from the filter paper blood spots using QIAamp DNA mini kit (Qiagen, Germany) as per manufacturer’s instructions and stored at –20°C for further use. A nested PCR was done for the PVMSP–3α gene as described by Manamperi et al., in 2008, with minor modifications (Manamperi et al., 2008). RFLP analysis of PVMSP–3α nested PCR products was carried out with Hha I restriction enzyme for further analysis of diversity (Hajime et al., 2011). Agarose gel electrophoresis was performed to detect all the amplified and digested DNA fragments.
Of the 26 samples 24 were amplified with MSP3α primers and two samples did not give any amplification. Based on the length of PCR products, four different allelic types of pvmsp–3α have been characterized around the world: type I (~1.9 Kb), type II (~1.1 Kb), type III (~1.5 Kb), and type IV (~0.3 Kb) (Huang et al., 2014). In the present study only type I and II were detected with higher prevalence of type II (59%) (Figure 1). RFLP analysis of the nested PCR product of PVMSP3α with restriction enzyme Hha I showed a size conservation of the largest fragment (approximately 1000 bp), while the size variation of smaller fragments show the presence of six different alleles in the studied cohort of isolates (Table 1).
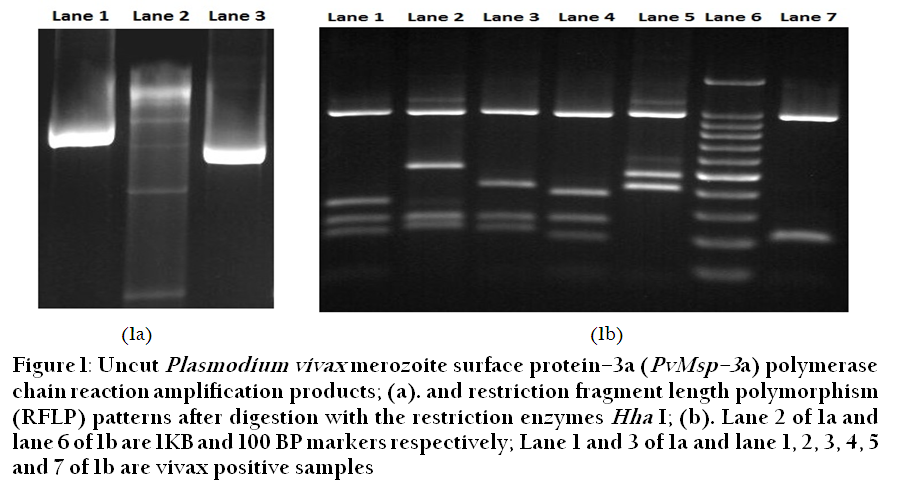
Figure 1: Uncut Plasmodium vivax merozoite surface protein–3a (PvMsp–3a) polymerase chain reaction amplification products
The present study demonstrates that the level of genetic diversity of the P.vivax population in Sri Lanka is remarkably lower than the levels in other tropical and subtropical areas reported by Prajapati et al., Adhikari et al., and Ribeiro et al., (Prajapati et al., 2010; Adhikari et al., 2012; Ribeiro et al., 2011). A study done in Chennai, India in 2010 showed 14 distinct alleles among 22 isolates (Prajapati et al., 2010). The studies done in Jhapa district in Nepal and in Brazil showed 13 in 56 isolates and 11 distinct alleles in 52 isolates respectively (Adhikari et al., 2012, Ribeiro et al., 2011).
A dramatic reduction of P. vivax case load has been observed in Sri Lanka over the period 2000–2011. The case load declined from 150,389 in the year 2000 to 1506 cases in 2005 and a mere 119 cases in 2011, with no indigenous cases being reported since October 2012. In order to determine the way that the population structure of P. vivax has changed during the last few years, the results of the present study was compared with two previous studies which also reported variations of PVMSP3α of Sri Lankan field isolates during the years 2000 and 2005. The study which was carried out from December 1998 to March 2000 analysed 196 P.vivax cases (0.1 % of the total P.vivax infected cases during the period of sample collection) and reported 3 major types with undigested PCR products and 22 different alleles after the RFLP yielding a highly diverse population of P.vivax (Wickramarachchi et al., 2010).
Another study which was done using 311 (5.93 % of the total infected cases) Sri Lankan P.vivax isolates collected from 2004 to 2006 also reported 3 major types after performing nested PCR and 11 different alleles with RFLP of Hha I digestion (Schousboe et al., 2011). Subsequently it was found that all 6 alleles found in the present study had been described within the 11 different alleles (Figure 2) of the previous study (Schousboe et al., 2011). Therefore it is also observed that there is a considerable genetic conservation in MSP3α among the P.vivax isolates in Sri Lanka over the last few years. In comparison to other studies the percentage of analysed cases (21.84 %) out of the total recorded vivax cases of this study was relatively large. Therefore it could be concluded that the level of genetic diversity in year 2011 is low even though the total number of samples analysed was small compared to the other two studies. Furthermore it is observed that the variation of PVMSP3α has been gradually undergoing reduction since year 2000.
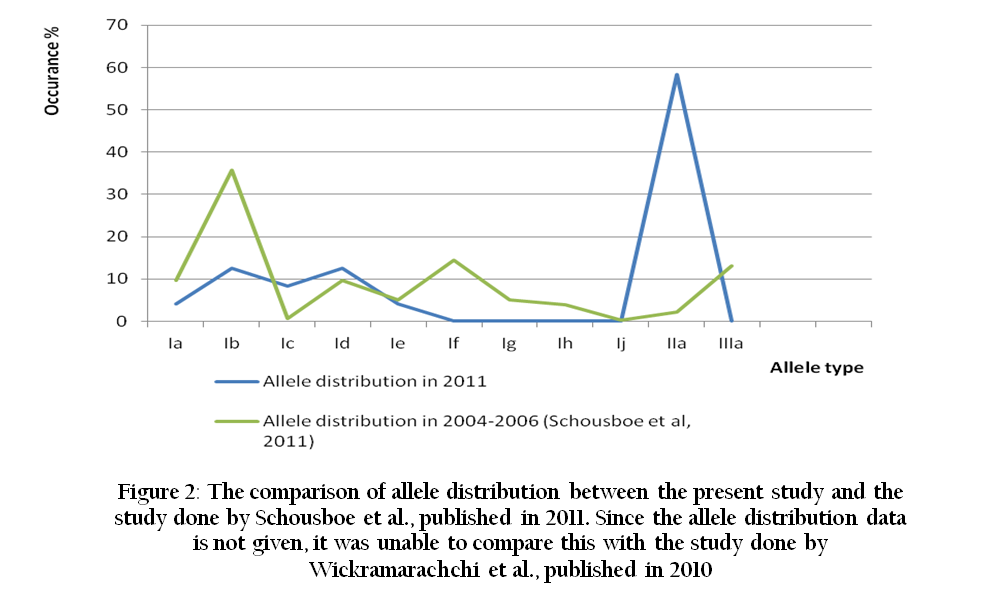
Figure 2: The comparison of allele distribution between the present study and the study done by Schousboe et al., published in 2011. Since the allele distribution data is not given, it was unable to compare this with the study done by Wickramarachchi et al., published in 2010
In the case of P. falciparum populations, the levels of genetic diversity are normally associated with the levels of malaria endemicity. That is, the levels of genetic diversity of the parasite populations are higher in high transmission areas and lower in low transmission areas, although some exceptions have been reported (Wagami et al., 2012). In contrast this is still not clearly understood in the case of P.vivax populations even though it is suggested that there may be some association between these two factors. Early gametocytogenesis may enhance the efficiency of transmission to Anopheles mosquitoes, thus allowing transmission to occur before symptoms appear. Thereby, the levels of genetic diversity tend to be high in P. vivax populations even in low transmission areas (Imwong et al., 2005; Ferreira et al., 2007). However the present study shows that there is a positive correlation between the levels of genetic diversity and the levels of malaria endemicity in P.vivax populations in Sri Lanka, although some studies contradict this finding (Imwong et al., 2005; Ferreira et al., 2007).
Even though this study shows a low polymorphism with respect to the MSP3α, previous studies which were carried out over the same period of time based on microsatellite markers showed a high genetic polymorphism among Sri Lankan P.vivax population (Gunawardena et al., 2010). Microsatellite variations are mainly derived by strand slippage during mitotic replication of the parasites, which is considered to mutate at high rates (Gunawardena et al., 2010). Even though the variation in microsatellites is so high, it has been reveled that it is not directly affected by host immunity and exhibits variation in the absence of selective pressure (Koepfli et al., 2009). Hence microsatellites are less reliable than surface antigenic markers to evaluate the dynamics of parasite transmission against the host immune system. This is supported by Sri Lankan epidemiological data where there are no indigenous vivax cases being reported since end of 2012 in the presence of high and low polymorphism with respect to the microsatellites and MSP3α respectively.
In conclusion, the transmission dynamics of P.vivax in Sri Lanka is being reduced parallel to the level of genetic diversity of PVMSP3α for the last few years. Therefore it could be suggested that the low level of diversity of P.vivax population may be a contributing factor towards elimination of malaria from the country as this can hinder the transmission ability of the parasite.
ACKNOWLEDGEMENT
We thank Dhammika N Gunasekara, Nandika Perera, Vajirapani de Silva and the staff of Genetech, and also to the Director and the staff of Anti Malaria Campaign, Sri Lanka. The study was financially supported by The Global Fund to Fight AIDS, Tuberculosis and Malaria (GFATM) under the grant number SRL–809–G12–M. The authors are grateful to the Director and the staff of GFATM at the Lanka Jatika Sarvodaya Shramadana Sangamaya, Sri Lanka.
REFERENCES
Adhikari M, Ranjitkar S, Schousboe ML, Alifrangis M, Imwong M, Bhatta DR, Banjara MR (2012). Genetic diversity of Plasmodium vivax merozoite surface protein –3α gene in Jhapa district of Napal. Southeast Asian J. Trop. Med. Public Health. 43: 2.
Anti Malaria Campaign (2011). annual report of the anti Malaria Campaign, 2011. Ministry of Health, Sri Lanka.
Ferreira MU, Karunaweera ND, Silva–Nunes M, Silva NS, Wirth DF (2007). Population structure and transmission dynamics of Plasmodium vivax in rural Amazonia. J. Infect. Dis. 195: 1218 – 1226.
http://dx.doi.org/10.1086/512685
PMid:17357061
Gunawardena S, Karunaweera ND, Ferreira MU, Phone–Kyaw M, Pollack RJ, Alifrangis M, Rajakaruna RS, Konradsen F, Amerasinghe PH, Schousboe ML, Galappaththy GNL, Abeyasinghe RR, Hartl DL, Dyann F (2010). Geographic Structure of Plasmodium vivax: Microsatellite Analysis of Parasite Populations from Sri Lanka, Myanmar, and Ethiopia. Am. J. Trop. Med. Hyg. 82(2): 235 – 242.
http://dx.doi.org/10.4269/ajtmh.2010.09-0588
PMid:20133999 PMCid:PMC2813164
Hajime H, Jung–Yeon K, Nirianne MQ, Toshihiro M, Wonja L, Toshihiro H, Kazuyuki T (2011). Recent increase of genetic diversity in Plasmodium vivax population in the Republic of Korea. Malar. J. 10: 257.
http://dx.doi.org/10.1186/1475-2875-10-257
PMid:21899730 PMCid:PMC3176257
Huang B, Huang S, Xin–zhuan S, Guo H, Xu Y, Xu F, Hu X, Yang Y, Wang S, Lu F (2014). Genetic diversity of Plasmodium vivax population in Anhui province of China. Malar. J. 13: 13.
http://dx.doi.org/10.1186/1475-2875-13-13
PMid:24401153 PMCid:PMC3893497
Imwong M, Pukrittayakamee S, Grüner AC, Rénia L, Letourneur F, Looareesuwan S, White NJ, Snounou G (2005). Practical PCR genotyping protocols for Plasmodium vivax using Pvcs and Pvmsp1. Malar. J. 4: 20. doi:10.1186/1475–2875–4–20.
http://dx.doi.org/10.1186/1475-2875-4-20
Koepfli C, Mueller I, Marfurt J, Goroti M, Sie A, Oa O, Genton B, Hans–Peter B, Felger I (2009). Evaluation of Plasmodium vivax Genotyping Markers for Molecular Monitoring in Clinical Trials. J. Infect. Dis. 199: 1074 – 80.
http://dx.doi.org/10.1086/597303
PMid:19275476
Manamperi A, Mahawithanage S, Fernando D, Wickremasinghe R, Bandara A, Hapuarachchi C, Abeyewickreme W, Wickremasinghe R (2008). Genotyping of Plasmodium vivax infections in Sri Lanka using Pvmsp–3α and Pvcs genes as markers: A preliminary report. Asian Pac J Trop Biomed. 25(2): 100 – 106.
Prajapati SK, Joshi H, Valecha N (2010). Plasmodium vivax merozoite surface protein–3α: a high–resolution marker for genetic diversity studies. J Vector Borne Dis. 47: 85 – 90.
PMid:20539045
Ribeiro RS, Ladeira L, Rezende AM, Fontes CJ, Carvalho LH, Ferreira C, Brito AD (2011). Analysis of the genetic variability of PvMSP–3α among Plasmodium vivax in Brazilian field isolates. Mem. Inst. Oswaldo Cruz. 106: 27 – 33.
http://dx.doi.org/10.1590/S0074-02762011000900004
PMid:21881754
Schousboe L, Rajakaruna S, Amerasinghe H, Konradsen F, Ord R, Pearce R, Bygbjerg C, Roper C, Alifrangis M (2011). Analysis of Polymorphisms in the Merozoite Surface Protein –3α Gene and Two Microsatellite Loci in Sri Lankan Plasmodium vivax: Evidence of Population Substructure in Sri Lanka. Am. J. Trop. Med. Hyg. 85(6): 994 – 1001.
http://dx.doi.org/10.4269/ajtmh.2011.11-0338
PMid:22144433 PMCid:PMC3225177
Wagami M, Fukumoto M, Hwang SY, Kim SH, Kho WG, Kano S (2012). Population Structure and Transmission Dynamics of Plasmodium vivax in the Republic of Korea Based on Microsatellite DNA Analysis. PLoS Negl. Trop. Dis. 6(4): 1592.
http://dx.doi.org/10.1371/journal.pntd.0001592
PMid:22509416 PMCid:PMC3317904
Wickramarachchi T, Premaratne PH, Dias S, Handunnetti SM, Randeniya PV (2010). Genetic complexity of Plasmodium vivax infections in Sri Lanka, as reflected at the merozoite–surface–protein–3α locus. Ann Trop Med Parasitol. 104(2): 95 – 108.
http://dx.doi.org/10.1179/136485910X12607012374190
PMid:20406577
WHO Global Malaria Programme. World Malaria report (2013). World Health Organization, 2013.


