
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 1 (4S): 24 – 29Special Issue–4 (Progress in Research on Viruses and Viral Diseases)
Isolation and Molecular Characterization of Bluetongue Virus 16 of Goat Origin from India
Abidali Ismailbhai Dadawala1, Hemendra Singh Kher1, Bharat Singh Chandel1, Abidali Gulamhaider Bhagat1, Harshad Chaturbhai Chauhan1, Koushlesh Ranjan2, Prasad Minakshi2*
- College of Veterinary Science and AH, Sardarkrushinagar Dantiwada Agricultural University Sardarkrushinagar, Gujarat, India, 385506
- Department of Animal Biotechnology, Lala Lajpat Rai University of Veterinary and Animal Sciences, Hisar, Haryana, India, 125004
*Corresponding author:minakshi.abt@gmail.com
ARTICLE CITATION:
Dadawala AI, Kher HS, Chandel BS, Bhagat AG, Chauhan HC, Ranjan K and Minakshi P (2013). Isolation and molecular characterization of bluetongue virus 16 of goat origin from India. Adv. Anim. Vet. Sci. 1 (4S): 24 – 29.
Received: 2013–09–10, Revised: 2013–11–15, Accepted: 2013–11–16
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/136/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Bluetongue (BT), transmitted by Culicoides (biting midges), is a disease of domestic and wild ruminants, infecting primarily sheep. The causative agent is bluetongue virus (BTV) which is double stranded RNA virus of the genus Orbivirus and family Reoviridae. A total of 38 blood samples from goat were screened for BTV using s–ELISA. Of the samples analyzed, only 12 were found positive. The Sandwich–enzyme linked immunosorbent assay (s–ELISA) positive samples were subjected to virus isolation in chicken embryo and in BHK–21 cell culture and only 5 samples were adapted to cell culture. The viral nucleic acid was extracted and BTV specific ten segmented genome with characteristics migration pattern of 3:3:3:1 was observed via RNA–PAGE (Polyacrylamide gel electrophoresis). BTV was further confirmed with group specific NS1 gene and VP2 gene based serotyping. All the five isolates were typed as BTV16. Phylogenetic analysis revealed that the isolates in this study clustered closely with BTV16 Indian isolates and were related to BTV16 isolates from Japan and Greece. The analysis revealed nucleotide (98.5%) and deduced amino acid sequence identity (97.5%) among themselves and with other BTV16 isolates reported previously from India. They also showed nucleotide (96.5 to 97.6%) and amino acid identity (96 to 98%) with BTV16 from Japan and Greece. The least identity of nucleotide (<93.1%) and amino acid (<98%) was observed with BTV16 isolate (DPP965) from Australia.
INTRODUCTION
Bluetongue (BT) is an infectious but non–contagious arthropod borne viral disease of domestic and wild ruminants belong to Genus Orbivirus and family Reoviridae. Bluetongue virus (BTV) causes economically important hemorrhagic disease mainly in sheep and occasionally in cattle and other species (Tabachnick, 2004). BTV is transmitted biologically by Haematophagous midges in the genus Culicoides, and infection occurs throughout the temperate regions (45 to 53 °N and 35 °S) of the world coincident with the distribution of competent Culicoides vectors (Gibbs and Greiner, 1994).
The disease has been included in the multi–species list of notifiable disease by Office International des Epizooties (OIE), Paris (OIE, 2013). It is one of the major barriers in International Trade of animals and its products. The economic losses due to BT are attributed to high morbidity, mortality, abortion, foetal death and deformities as well as milk, meat and fleece losses (Tatem et al., 2003). Therefore, the mandatory export restrictions and the surveillance requirements are imposed on movement of ruminant animals and their products of BT endemic countries to BT free countries (Velthuis et al., 2009).
Most common symptoms of BT disease are high fever with excessive salivation, swollen lips and tongue, petechial haemorrahage, congestion and small ulcers on the mucous membranes of the mouth and conjunctiva, coronitis and reproductive disorders leading to abortion or congenital deformities (Anderson and Jensen, 1969; Ataseven et al., 2006). Clinical BT has been observed in sheep, but clinical form has not been noticed in goats, in spite of high percentage of BT seroprevalance in goats. Cattle mainly act as reservoir host (Luedke and Jones 1984, Mehrotra, et al. 2001).
The BTV genome contains ten segmented dsRNA, which encodes seven structural proteins VP1 to VP7 and four non–structural proteins viz. NS1, NS2, NS3/NS3a and NS4 (Mertens et al., 1989; Ratinier et al., 2011). The twenty four distinct BTV serotypes of BTV have been reported globally (Martens et al., 2005). However, recently two more serotypes BTV25 from Switzerland (Hofmann et al., 2008) and BTV26 from Kuwait (Maan et al., 2011) have been reported. Being India the endemic for BT, the 21 different serotypes have been reported from Indian subcontinent (Prasad et al., 2009). BT is endemic in many states of India. During monsoon season of 1997 – 98 it caused death of 300,000 sheep and goats in Tamil Nadu state (Ilango, 2006). Recently, BTV serotype 21 has been reported from west Bengal state (Jordar et al., 2009). Seroprevalance of 15 distinct BTV serotypes have been reported from Gujarat state (Chandel et al., 2004). The BTV16 from sheep origin was reported earlier in India (Shafiq et al., 2013). The complete genome sequence of BTV16 of goat origin from southern part of India has been done recently (Minakshi et al., 2012). However, the little information is available regarding the BTV from goat origin in Gujrat state of India. Therefore, the present investigation was carried out to serotype identification of BTV of goat origin using vp2, serotype specific RT–PCR and nucleic acid sequencing of segment 2. The sequence data was compared with sequences of similar serotypes from India and other countries from throughout the world.
MATERIALS AND METHODS
Sample Preparation and Virus Isolation
The blood samples were collected from goats reared at Sheep and Goat Unit, Sardarkrushinagar, (having previous history of BTV outbreak) showing clinical signs suggestive of Bluetongue (BT) or having previous history of abortion. Subsequently, a total of 38 blood samples were screened for detection of BTV antigen using s–ELISA kit (IVRI, Mukteswar). The s–ELISA positive samples were processed for virus isolation in 6 day old embryonated chicken embryo (ECE) and subsequently adapted in BHK–21 cell lines. The heparinised blood samples were subjected to centrifugation at 800×g for 10 minutes in refrigerated centrifuge. The pelleted erythrocytes obtained were washed thrice in CMF–PBS buffer and sonicated at 15 mA current for 30 seconds under cold condition by Ultrasoniocator (Labsonic P, Sartorius). After sonication, the samples were filtered through membrane filter (0.22 m) and the filtrates were used as inoculum for virus isolation.
The each sample was inoculated in five different 6–day–old ECE using 0.1 mL of inoculum via yolk–sac (YS) route and incubated at 33.5 oC for seven days. The whole infected embryos were harvested, decapitated, pooled and finally homogenized in CMF–PBS buffer to get 10 percent (w/v) of suspension. After three cycles of freeze–thaw, the homogenized suspension was centrifuged and supernatant was used for next passage. A total of four such passages were given into chick embryos. After the fourth passage, the chick embryo suspension was diluted further so as to get 10 per cent (w/v) suspension for virus isolation in cell culture. Five hundred microlitres of 10 percent embryo suspension obtained from ECE were inoculated on to confluent BHK–21 cell monolayer. The inoculated BHK–21 cells were incubated at 37oC. After three passages in BHK–21 cell culture BTV specific visible cytopathic effect (CPE) such as rounding, detachment and foamy degeneration of cells were observed.
Viral Nucleic Acid Extraction
A total of five samples (G4/IND/2011, G10/IND/2011, G52/IND/2011, G53/IND/2011 and G83/IND/2011) showing BTV specific CPE were further grown in BHK–21 cell line up to six passages. The viral nucleic acid (dsRNA) was extracted from BHK–21 cells grown viruses using modified Trizol method (Chomoczynski and Sacchi, 1987).
RNA Poly acrylamide Gel Electrophoresis of Viral Nucleic Acid (RNA–PAGE)
The dsRNA nucleic acid was subjected for 8% poly acrylamide gel electrophoresis to determine the specific pattern of BTV specific nucleic acid. The RNA–PAGE was allowed for silver staining to visualize the nucleic acid segments (Svensson et al., 1986) (data not shown).
cDNA Preparation and PCR
Viral genomic dsRNA was used as template for cDNA synthesis using random decamer primer (Ambion, USA) in thermal cycler (Biorad i Cycler). The reverse transcription reaction was carried out using Mo–MuLV reverse transcriptase (Sibzyme) as per manufacturer’s instructions.
The cDNA obtained was subjected to PCR using BTV group specific ns1 gene primers P1: 5’GTT CTC TAG TTG GCA ACC ACC3’ and P2: 5’ AAG CCA GAC TGT TTC CCG AT3’ to generate an amplicon of 274bp size (Prasad et al., 1999). The NS1 gene based group specific PCR confirmed the samples as BTV.
Serotype Confirmation and Nucleic Acid Sequencing
BTV isolates confirmed by RNA–PAGE and NS1 gene based RT–PCR were submitted to Bluetongue virus serotyping centre, AINP–BT, Department of Animal Biotechnology, Lala Lajpat Rai University of Veterinary and Animal Sciences, Hisar for identification of serotypes. The serotypes of virus isolates were confirmed by VP2 gene based type specific PCR using all the 24 different BTV serotype specific primers and nucleic acid sequencing. The serotype specific PCR products of VP2 gene were sequenced in the automatic DNA sequencer AB1 PRISMTM 3100 version 3.0.
Sequence Data Analysis
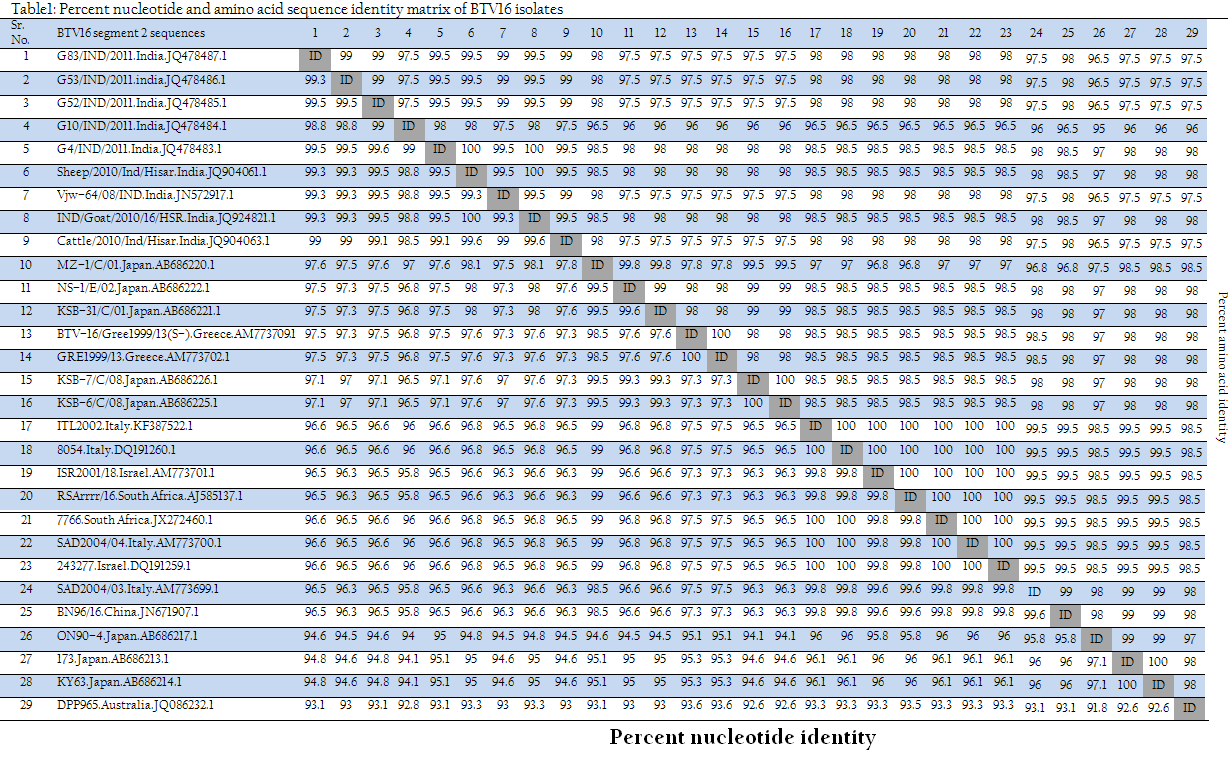
The VP2 gene sequences of all the isolates obtained were subjected to BLASTN+ 2.2.28 (Zhang et al., 2000) analysis for sequence based serotype confirmation. The sequence data obtained was submitted to NCBI and accession number for isolates G4/IND/2011, G10/IND/2011, G52/IND/2011, G53/IND/2011 and G83/IND/2011 was assigned as JQ478483.1, JQ478484.1, JQ478485.1, JQ478486.1 and JQ478487.1, respectively. The percent nucleotide and its deduced amino acid sequence identity matrix with global isolates were calculated using BioEdit 7.2.3 (Hall, 1999). The phylogenetic analysis of VP2 gene sequences used in this study along with other global sequences of same serotype was done using MEGA5 software (Tamura et al., 2007).
RESULT AND DISCUSSION
BTV is endemic in Indian subcontinent and a large number of BTV serotypes were reported from Gujarat state. In present study 38 blood samples of goat origin were initially screened using s–ELISA kit. Out of these samples, 12 had shown positive result for BTV. Recently, higher incidence (29.42 percent) of BTV from suspected field samples such as blood, washed RBCs, buffy coat and plasma from animals, using an in–house developed Mab based s– ELISA test has been reported (Gandhale et al., 2010).
The s–ELISA positive samples were further processed for virus isolation first in ECE followed by BHK–21 cell lines. Out of 12 samples 5 were adapted to cell culture. In chick embryo pathogenicity of all the isolates appeared to be on increasing trend with the higher passage level. In most of the cases, embryos died between third and fifth day post inoculation. The characteristic embryopathic changes in ECE suggestive of BTV such as cherry red discoloration, oedema, cutaneous hemorrhages and stunted growth were noticed in all the harvested embryos (Abu Elzein et al., 1992; Ramesh Babu et al., 1992; Saravanabava and Venkatesan 1995; Chandel 1996 and Mahdvi et al. 2006). Whereas, the control ECE of same age didnot show any such changes. The samples not showing any signs of viral pathogenicity upto 4th passage were considered negative.
Out of the 12 s–ELISA positive samples, 5 isolates viz. (G4/IND/2011, G10/IND/2011, G52/IND/2011, G53/IND/2011 and G83/IND/2011) were recovered in ECE and all isolates adapted well to BHK–21 cell lines and produced visible characteristic CPE specific to BTV from third passage and onwards. Periodical examination of infected BHK–21 monolayers showed initially rounding off and increase in number of floating cells in medium, followed by formation of empty areas in monolayers (Sekar et al., 2009).
In RNA–PAGE, all the five isolates showed the presence of classical 10 segments of dsRNA with 3:3:3:1 pattern characteristic to BTV (data not shown). All the isolates yielded PCR amplicon of expected size of 274bp in ns1 gene based RT–PCR (data not shown). The characteristics CPE in BHK–21 cell culture, migration pattern in RNA–PAGE and an expected PCR amplicon with ns1 gene RT–PCR confirmed the samples as BTV. Several other workers have evaluated different RT–PCR assays to study the BTV isolates grown in BHK–21 cell monolayers (Aradabib et al., 1994; 1995; 1998; Breard et al., 2003; Biswas et al., 2010).
For serotype confirmation, these isolates were submitted to BTV typing Centre, AINP–BT, Department of Animal Biotechnology, LLRUVAS, Hisar. All the isolates were subjected to vp2 gene specific RT–PCR using primers specific to vp2 gene of all the 24 BTV serotypes. All the isolates in study had shown specific PCR amplicon of 768bp with BTV16 vp2 gene primer only (Figure 1). However, they did not give any amplification with remaining serotype specific primers. Thus, all the isolates were typed as BTV16. Furthermore, vp2 gene specific PCR products of all six isolates were directly sequenced in the automatic DNA sequencer for serotype confirmation. BLASTN+ 2.2.28 analysis of all the isolates showed that segment 2 (VP2 gene) of all the isolates align only with BTV16 isolates from different parts of the world, confirming all the isolates as BTV16. The percent nucleotide and its deduced amino acid sequences identity of the isolates in study along with other BTV16 isolates from different parts of the world were calculated using Bioedit 7.2.3 (Hall, 1999).

The sequence identity revealed that the isolates in study along with other BTV16 isolates from India showed more than 98.5% nucleotide and more than 97.5% deduced amino acid identity (Table 1). However, they also showed higher identity (nucleotide 96.5 to 97.6% and amino acid 96 to 98%) with BTV16 from Japan (NS–1/E/02, KSB–31/C/01, KSB–7/C/08, KSB–6/C/08 and MZ–1/C/01) and Greece (BTV–16/Gree1999/13(S–2), GRE1999/13) (Table 1). They showed less than 96.6% nucleotide and less than 98.5% amino acid identity with various isolates from Italy, Israel, South Africa, Japan and China. However, these isolates showed least nucleotide and amino acid identity of <93.1% and <98% respectively with BTV16 isolate (DPP965) from Australia.
Phylogenetic analysis of these isolates along with other global isolates using Mega 5 software programme revealed that the isolates in present study formed a separate closer cluster with BTV16 Indian isolates. They were also closely related to BTV16 isolates from Japan and Greece. However, they were distantly related to cluster of Italy, Israel, South Africa and China and Japan. Australian BTV16 isolate (DPP965) represent a separate cluster and was distantly related to isolates in study (Figure 2). However, to know the origin of individual segments of the genome, sequencing of the all the segments is required. Recently, reassortment in the segment 6 of BTV16 and BTV21 has been reported in the Indian BTV16 isolate (Shafiq et al., 2013).
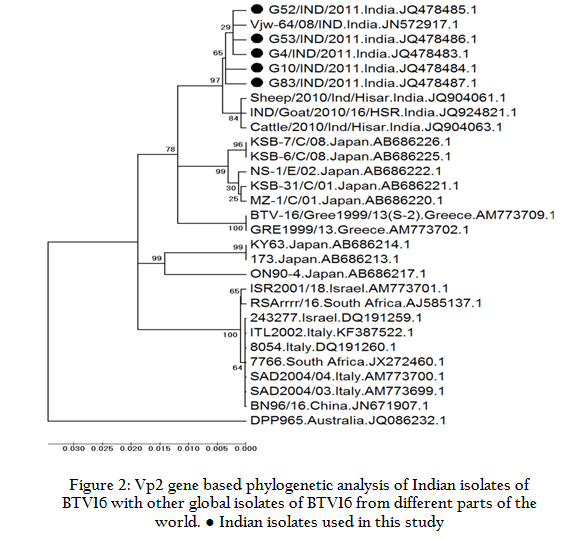
Figure 2: Vp2 gene based phylogenetic analysis of Indian isolates of BTV16 with other global isolates of BTV16 from different parts of the world. ● Indian isolates used in this study
The sequence identity and phylogenetic analysis revealed that the BTV16 isolates in study might be originated either from Indian or Greece or Japanese BTV16 isolates. The transmission of BTV takes place along with live animals or its product transport/movement and its Culicoides vector. The potential BTV vector, Culicoides Oxystoma is prevalent in entire south East Asian region including India and Japan, which is responsible for transmission of BTV in these regions along with live animal movement.
There are 26 recognized serotypes of BTV prevalent throughout the world (Maan et al., 2011), which may be due to the segmented genome of the virus and continuous genetic reassortment occurring in nature. The genetic diversity and common antigens among various BTV serotypes and other Orbiviruses make control of this disease difficult. Moreover, the virus is transmitted by Culicoides vectors, posing the disease control a challenge.
Therefore, to device effective protective measures, knowledge about the prevalence of different BTV serotypes and diversity within and between them is of fundamental importance. The serotype–specific RT–PCR is a rapid assay that has shown perfect agreement with the serotyping by conventional virus neutralization methods (Mertens et al., 2007). Rapid molecular based diagnosis of BTV isolates is required to improve surveillance of BTV serotypes prevalent and to facilitate the choice of an appropriate serotype–specific vaccine in a disease outbreak. In the present study, six isolates from goat blood were typed as BTV–16. This appears to be a first report in isolation of BTV from Goats in Gujarat and only second in the country. This study has attested for the need of inclusion of BTV–16 in the "Multivalent vaccine” which is presently under development/trial against BTV.
CONCLUSION
BT is one of the major infectious diseases of small ruminants in India. India is endemic for BT disease and 22 different BTV serotypes were reported from this country. BT is primarily a disease of sheep. In this paper we have reported BT in goat. All the positive samples for BTV were confirmed as BTV16 based on segment 2 RT–PCR and nucleotide sequencing. The sequence and phylogenetic analysis revealed that all the five isolates might be originated either from other Indian BTV16 isolates or from BTV16 from Japan or Greece. The BTV16 is one of the most prevalent serotype in India. The segment 2 based RT–PCR used here can be used for diagnosis of BTV from variety of biological samples such as blood, tissue samples and Culicoides vectors.
ACKNOWLEDGEMENT
Authors acknowledge the support received from, SDAU, Dean, College of veterinary Science and AH and In–charge, Sheep and Goat Unit, SDAU, Sardarkrushinagar. This work has been supported by ICAR All India Network Project on Bluetongue.
CONFLICT OF INTEREST
We declare that we don’t have any conflict of interest.
REFERENCES
AbuElzein EME, Gameel AA, Al–Afaleg AI and Hassanein MM (1992). Isolation of a virus serologically related to the bluetongue group from an outbreak of haemorrhagic disease among deer in Saudi Arabia. Vet. Rec. 7: 439 – 441.
http://dx.doi.org/10.1136/vr.131.19.439
Anderson CK and Jensen R (1969). Pathologic changes in placentas of ewes inoculated with bluetongue virus. Am. J. Vet. Res. 30: 987 – 999.
PMid:4307344
Aradaib IE, Akita GY and Osburn BI (1994). Detection of epizootic hemorrhagic disease virus serotypes 1 and 2 in cell culture and clinical samples using polymerase chain reaction. J. Vet. Diagn. Invest. 6:143 – 147.
http://dx.doi.org/10.1177/104063879400600202
PMid:8068743
Aradaib IE, Akita GY, Pearson JE and Osburn BI (1995). Comparison of polymerase chain reaction and virus isolation for detection of epizootic hemorrhagic disease virus in clinical samples from naturally infected deer. J. Vet. Diagn. Invest. 7:196 – 200.
http://dx.doi.org/10.1177/104063879500700205
PMid:7619901
Aradaib IE, Schore CE, Cullor JS and Osburn BI (1998). A nested PCR for detection of North American isolates of BTV based on NS1 genome sequence analysis of BTV–17. Vet. Microbiol. 59: 99 – 108.
http://dx.doi.org/10.1016/S0378-1135(97)00176-4
Ataseven VS, Ataseven L, Tan T, Babür C and Oguzoglu CT (2006). Seropositivity of agents causing abortion in local goat breeds in Eastern and South–eastern Anatolia, Turkey. Revue Méd Vét. 157: 545 – 550.
Biswas SK, Chand K, De A, Pandey LK, Mohapatra JK, Prasad G, Mondal B (2010). Isolation of bluetongue virus serotype 1 (BTV–1) from goats and its phylogenetic relationship with other BTV–1 isolates worldwidebased on full–length sequence of genome segment–2. Archives of Virol. 155:2041 – 2046.
http://dx.doi.org/10.1007/s00705-010-0794-x
PMid:20842392
Bréard E, Sailleau C, Coupier H, Mure–Ravaud K, Hammoumi S, Gicquel B, Hamblin C, Dubourget P. and Zientara S. (2003). Comparison of genome segments 2, 7 and 10 of bluetongue virus serotype 2 for differentiation between field isolates and the vaccine strain. Vet. Res. 34:777 – 789.
http://dx.doi.org/10.1051/vetres:2003036
PMid:14746772
Chandel BS (1996). Seroepidemiology, isolation and pathogenicity of Bluetongue virus. Ph.D. Thesis submitted to Gujarat Agricultural University, Sardarkrushinagar.
Chandel BS, Kher HN, Chauhan HC and Vasava KA (2004). Serological survey of antibodies to Bluetongue virus in domestic ruminant in Gujarat. Indian Vet J. 81:737 – 740.
Chomoczynski P and Sacchi N (1987). Single step method of RNA isolation by acid Guanidinium isothiocyanate – phenol – chloroform extraction. Analit. Biochem. 162:156 – 159.
http://dx.doi.org/10.1006/abio.1987.9999
http://dx.doi.org/10.1016/0003-2697(87)90021-2
Gandhale PN, Bhanuprakash V, Balamurugan V, Hosamani M, Venkatesan G and Singh RK (2010). Detection of bluetongue virus group–specific antigen using monoclonal antibody based sandwich ELISA. Virol. Sin. 25:390 – 400.
http://dx.doi.org/10.1007/s12250-010-3160-y
PMid:21221917
Gibbs EP and Greiner EC (1994). The epidemiology of bluetongue. Comp. Immunol. Microbiol. Infect. Dis.17:207–20.
http://dx.doi.org/10.1016/0147-9571(94)90044-2
Hall TA. (1999). BioEdit: a user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 41:95 – 98.
Hofmann, M.A., Renzullo, S., Mader, M., Chaignat, V., Worwa, G. and Thuer, B. (2008). Genetic Characterization of Toggenburg Orbivirus, a New Bluetongue Virus, from Goats, Switzerland. Emerg. Infect. Dis. 14: 1855 – 1861.
http://dx.doi.org/10.3201/eid1412.080818
PMid:19046507 PMCid:PMC2634640
Ilango K. (2006). Bluetongue virus outbreak in Tamil Nadu, southern India: Need to study the Indian biting midge vectors, Culicoides Latreille (Diptera: Ceratopogonidae). Current Science. 90:163 – 167.
Jordar SN, Lodh C, Chakrabarti A, Baksi S and Aich R. (2009). Isolation of Bluetongue virus serotype 15 and 21 in West Bengal, India. Vet. Rec. 165:751 – 752.
Luedke AJ and Jones RH (1984). Bluetongue diagnosis and significance in bovine animal. Bovine Practice. 15:70 – 86.
Maan S, Maan NS, Nomikou K. Batten C, Antony F, Belaganahalli MN, Attia MS, Reda AA, Al–Rashid SA, El Batel M, Oura CAL and Mertens PPC (2011). Novel Bluetongue Virus Serotype from Kuwait. Emerg. Infect. Dis. 17:886 – 889.
http://dx.doi.org/10.3201/eid1705.101742
PMid:21529403 PMCid:PMC3321788
Mahdavi S, Khedmati K and PishraftSabet L (2006). Serologic evidence of bluetongue infection in one humped camels (camelusdromedarius) in Kerman province, Iran. Iranian J. Vet. Res. 7:85 – 87.
Mehrotra ML, Ramakrishnan MA, Tembhurne PA and Pandey AB (2001). Immunogenic response of some Indian isolates of bluetongue virus. Presented at Seminar–cum–Workshop on "Bluetongue virus epidemiology and laboratory diagnosis", held at Punjab Agri. University, Ludhiana during February 19 – 22, 2001.
Mertens PP, Pedley S, Cowley J, Burroughs JN, Corteyn AH, Jeggo MH, Jennings DM, Gorman BM. (1989). Analysis of the roles of bluetongue virus outer capsid proteins VP2 and VP5 in determination of virus serotype. Virol. 170:561 –565.
http://dx.doi.org/10.1016/0042-6822(89)90447-9
Mertens PPC, Maan S, Samuel A and Attoui H (2005). Orbivirus Reoviridae. In: Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA, eds. Virus. Taxonomy VIIIth Report of the ICTV Elsevier/Academic Press London. Pp 466 – 483.
Mertens PPC,Maan NS, Prasad G, Samuel AR, Shaw A E, Potgieter AC, Anthony SJ and Maan S (2007). Design of primers and use of RT–PCR assays for typing European bluetongue virus isolates: differentiation of field and vaccine strains. J. Gen. Virol. 88:2811 – 2823.
http://dx.doi.org/10.1099/vir.0.83023-0
PMid:17872535
Minakshi P, Singh R, Ranjan K, Kumar P, Joshi CG, YKM Reddy and Prasad, G. (2012). Complete genome sequence of bluetongue virus serotype 16 of goat origin from India. J. Virol. 86: 8337 –8338.
http://dx.doi.org/10.1128/JVI.01128-12
PMid:22787269 PMCid:PMC3421637
OIE (2013). OIE–Listed diseases, infections and infestations in force in 2013. http://www.oie.int/animal-health-in-the-world/oie-listed-diseases-2013/.
Prasad G, Minakshi, Malik Y and Maan S (1999). RT PCR and its detection limit for cell culture grown bluetongue virus1 using NSI gene specific primers. Indian J. Exp. Biol. 37:1255 – 1258.
PMid:10865895
Prasad G, Sreenivasulu D, Singh KP, Mertens PPC and Maan S (2009). Bluetongue in the Indian subcontinent. In: Bluetongue. (Eds. Mellor P, Baylis M and Merten P C). Elsevier Ltd., London. 167 – 195.
http://dx.doi.org/10.1016/B978-012369368-6.50012-5
PMid:19339108
Ramesh Babu, N.G., Byregowda, S.M., Bragitha, A.J. and Gopal, T. (1992). Isolation and preliminary characterization of bluetongue virus from sheep. Indian Vet. J. 69:1071 – 1074.
Ratinier M, Caporale M, Golder M, Franzoni G, Allan K, Armezzani A, Bayoumy A, Rixon F, Shaw A, Palmarini M (2011). Identification and Characterization of a Novel Non–Structural Protein of Bluetongue Virus. PLoS Pathog. 7: e1002477. doi:10.1371/journal.ppat.1002477.
http://dx.doi.org/10.1371/journal.ppat.1002477
Saravanabava K and Venkatesan RN (1995). Epizootiology of bluetongue in sheep. Indian J. Anim. Sci. 65:246 – 249.
Sekar P, Ponmurugan K and Gurusubramanian G (2009). Comparative Susceptibility of BHK 21 and Vero Cell Lines to Bluetongue Virus (BTV) Isolate Pathogenic for Sheep. Internet. J. Microbiol. 7 (1):1 – 5.
Shafiq M, Minakshi P, Bhateja A, Ranjan K, Prasad G (2013). Evidence of genetic reassortment between Indian isolate of bluetongue virus serotype 21 (BTV–21) and bluetongue virus serotype 16 (BTV–16). Virus. Res. 173:336–43. doi: 10.1016/j.virusres.2013.01.009.
http://dx.doi.org/10.1016/j.virusres.2013.01.009
Svensson L, Uhnoo I, Grandien M and Wadeli G (1986). Molecular epidemiology of rotavirus infections in Upsala. Sweden. 1981; disappearance of a predominant electropherotype. J. Med. Virol. 18: 101 – 111.
http://dx.doi.org/10.1002/jmv.1890180202
PMid:3005484
Tabachnick WJ (2004). Culicoides and the global epidemiology of bluetongue virus infection. Vet. Ital. 40: 145 – 150.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, and Kumar S (2011). MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Bio. Evo. 28: 2731 – 2739.
http://dx.doi.org/10.1093/molbev/msr121
PMid:21546353 PMCid:PMC3203626
Tatem AJ, Baylis M, Mellor PS, Purse BV, Capela R, Pena I and Rogers DJ (2003). Prediction of bluetongue vector distribution in Europe and north Africa using satellite imagery. Vet. Microbiol. 97:13 – 29.
http://dx.doi.org/10.1016/j.vetmic.2003.08.009
PMid:14637035
Velthuis AG, Saatkamp HW, Mourits MC, de Koeijer AA, and Elbers AR (2009). Financial consequences of the Dutch bluetongue serotype 8 epidemics of 2006 and 2007. Prev. Vet. Med. 93: 294 – 304.
http://dx.doi.org/10.1016/j.prevetmed.2009.11.007
PMid:19962204
Zhang Z, Schwartz S, Wagner L and Miller W (2000). A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 7:203 –214
http://dx.doi.org/10.1089/10665270050081478
PMid:10890397

