
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 1 (5): 151 – 156Effect of Foot and Mouth Disease Vaccination on the Semen Quality of Mithun (Bos Frontalis)
Ponraj Perumal1, Kezhavituo Vupru2, Kobu Khate2, Muthusamy Veeraselvam3, Atul Kumar Verma4
- Scientist, Animal Reproduction Laboratory, National Research Centre on Mithun (ICAR), Jharnapani, Nagaland – 797 106, India
- Farm Manager, National Research Centre on Mithun (ICAR), Jharnapani, Nagaland – 797 106, India
- Assistant Professor, Veterinary University Training and Research Centre, TANUVAS, Salem, India
- Veterinary Officer, Sitapur, Department of Animal Husbandry and Veterinary Sciences, Government of Uttar Pradesh, India
*Corresponding author:perumalponraj@gmail.com
ARTICLE CITATION:
Perumal P, Vupru K, Khate K, Veeraselvam M and Verma AK (2013). Effect of foot and mouth disease vaccination on the semen quality of Mithun (Bos Frontalis). Adv. Anim. Vet. Sci. 1 (5): 151 – 156.
Received: 2013–09–07, Revised: 2013–09–23, Accepted: 2013–09–23
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/106/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
The present study was conducted to assess the effect of Foot and Mouth disease (FMD) vaccination on seminal parameters such as sperm motility, livability, total morphological abnormality, acrosomal and plasma membrane integrity and antioxidants profiles such as reduced glutathione (GSH), glutathione reductase (GRD), glutathione peroxidase (GPD), superoxide dismutase (SOD), catalase (CAT) and total antioxidant capacity (TAC) of semen of mithun breeding bulls, maintained at Semen Collection Centre, NRC on Mithun, Jharnapani, Nagaland. A total of 160 semen ejaculates of averaged 1.5 ml (1.39 ±0.59 ml) were collected from eight mithun bulls twice a week for five weeks before and five weeks after vaccination (Raksha–Ovac Trivalent vaccine) through rectal massage method and were used to assess the harmful effects through routine seminal and biochemical profile examination of semen. Results revealed that FMD vaccination affected the sperm functional parameters, antioxidant and biochemical profiles significantly (p<0.05) in mithun bulls. Similarly, white blood cell count of mithun was also affected significantly (p<0.05). The harmful effects of vaccination on these profiles suggested that the semen collection and preservation should be stopped upto restoration of normal fertility of sperm to avoid conception failure from artificial insemination using such semen in this precious species.
INTRODUCTION
Mithun (Bos frontalis) is a free–range bovine animal spread in the north–eastern hilly (NEH) region of India and is considered to be originated from wild Indian gaur (Simoons, 1984). According to livestock Census 2003, total mithun population in India is 2, 64, 279, of which, Arunachale Pradesh is home to 82.84% (2, 18, 931) of mithun followed by 12.63% (33, 385) in Nagaland, 3.79% (10, 024) in Manipur and 0.73% (1,939) in Mizoram. Mithun also plays an important role in the upliftment of socio–economic life of its rearers in this region (Mondal and Pal, 1999). The recent activities to enhance the population size as an economic beef animal and is fulfilled by its rearing under semi–intensive system and adoption of controlled breeding programme. In this context, it is important to standardize effective semen collection and preservation protocol for this mountain cattle to adopt controlled breeding and artificial insemination (AI) programme for breed improvement of this species.
In India, like other bovines species, mithuns are reared in semi–intensive system suffers from various infectious diseases like Tuberculosis, Para–tuberculosis, Brucellosis, Foot and Mouth disease (FMD), Bovine Viral Diarrhoea and Infectious Bovine Rhinotracheitis (IBR) (Rajkhowa et al., 2003), which has tremendous detrimental effect on profitable mithun husbandry practices. But mithun is affected severely with foot and mouth disease (Dutta et al., 1979; Verma and Sarma, 1997) with strain of Pan Asia I (OIE/FAO, 2010), O strain (Hegde et al., 2011) and type Asia –1 strain (Verma and Sarma, 1997) in this region of India. Frequent outbreak of FMD is also reported in mithun (Verma and Sarma, 1997; Barman et al., 1999).
The mithun breeding bulls are vaccinated against foot and mouth disease as a regular prophylactic measure. There are various reports of harmful effects of vaccination on the seminal parameters and its quality in cattle and buffalo (Mangurkar et al., 2000; Murugavel et al., 1997; Bhakat et al., 2010; Bhakat et al., 2008; Pankaj et al., 2007). But no proper report in mithun species. However, the available reports are conflicting about the quality of semen due to vaccination in cattle. Some of them reported that it does not have significant effect on seminal parameters and its quality (Mangurkar et al., 2000), whereas others were found that higher incidence of sperm abnormalities in the vaccinated semen (Kammar and Gangadhar, 1998). These seminal parameters and its quality may be affected by this FMD vaccination due to vaccine stress and anaphylactic shock (Murugavel et al., 1997) resulting from increased temperature of the body as well as testes. But the recovery rate of semen quality is depending upon the duration, nature of the thermal insult and type of the vaccine.
Mammalian sperm membrane has high polyunsatured fatty acids and sperms are very susceptible to ROS production and affects motility and plasma membrane, acrosome and DNA integrity (Griveau et al., 1995). The rate of generation of ROS in testis and/or spermatozoa depends upon the temperature in that (Ikeda et al., 1999) as vaccination leads to thermo stress causes deterioration of sperm quality. It is therefore crucial for the male reproductive system to be well guarded against oxidative injury. Mithun semen normally contains anti–oxidants, including GSH, SOD, CAT, GRD, GPD and TAC can offset lipid peroxidation (Perumal et al., 2013a; Perumal et al., 2013b). But the concentration of these antioxidants is reduced due to thermal stress and anaphylactic shock in vaccinated animals (Ahotupa and Huhtaniemi, 1992) as these antioxidants are derived from epididymis (Fouchecourt et al., 2000; Zini et al., 2002) and seminal vesicle (Tramer et al., 1998) into the semen as the epididymis and accessory sex glands are thermo sensitive and androgen dependent (Saeed et al., 1994). Moreover, the thermal stress affects the GSH oxidation and reduction cycle in spermatozoa and seminal plasma of mammalian semen (Ahotupa and Huhtaniemi, 1992). There was no report on harmful effects of vaccination on semen of mithun and to the best of our knowledge, this is the first report in this species. Therefore the present study was planned to assess the effects of FMD vaccination on seminal parameters and antioxidants profiles in mithun.
MATERIALS AND METHODS
Animals and Semen Collection
Eight apparently healthy mithun bulls of 4 to 6 years of age (5.13 ± 0.91) were selected randomly from the mithun herd of NEH region. The average body weight of the bulls from 501.25 ± 6.23 Kg (493 to 507 Kg) at 4 yr to 530.50 ± 7.59 Kg (523 to 538 Kg) at 6 yr of age with body condition of score 5–6 were maintained under uniform feeding, watering, housing, lighting conditions and management. Experimental animals were offered daily ad libitum drinking water, 30 kg mixed jungle forages containing 18.4% dry matter and 10.2% crude protein, 4 kg concentrates containing 87.1% dry matter and 14.5% crude protein fortified with mineral mixture as well as required salt. Semen ejaculates were collected by per rectal massage method (Karunakaran et al., 2007) twice a week before and after vaccination every week for 5 weeks, were used to study the harmful effects of vaccination. Trivalent vaccine containing virus serotypes O, A and Asia–1(Raksha–Ovac, Indian Immunological Limited, India) was administered by deep intra muscular injection route. These semen ejaculates were subjected to evaluation for individual motility, concentration by haemocytometer method (Tomar, 1997), livability by eosine–nigrosine stain (Robeck and O’Brien, 2004), acrosomal integrity by Giemsa stain (Watson, 1975) and plasma membrane integrity by hypo–osmotic swelling test (HOST) (Buckett et al., 1997) as per the standard procedure using microscope (Nikon, Eclipse 80i, Japan). The antioxidant profiles such GSH, GPD, GRD, SOD, CAT and TAC and biochemical profiles such as total cholesterol were estimated by commercial diagnostic kits (BioVision®, CA 95035, USA). LPO level of sperm and seminal plasma was measured by determining the malondialdehyde (MDA) production by using thiobarbituric acid (TBA) (Buege and Aust, 1978; Suleiman et al., 1996). The blood sample was collected immediately after collection of semen and counted the total white blood cells by standard method. During the present study, all the experimental protocols were approved by the Institute Animal Ethics Cell.
Statistical Analyses
The results of experiment were analysed statistically and expressed as the mean ± S.E.M. Means of seminal parameters such as sperm concentration, individual motility, livability, total sperm abnormality, plasma membrane and acrosomal integrity, antioxidant profiles such as GSH, GPD, GRD, CAT, SOD and TAC, biochemical profiles such as MDA production and cholesterol concentration and white blood cell count were analyzed again by student‘t’ test between the pre and post vaccination stages using the SPSS (version 15.0; SPSS, Chicago, IL). Differences with values of p < 0.05 were considered to be statistically significant after arcsine transformation of percentage data.
RESULTS
The effects of FMD vaccination on various seminal parameters, antioxidants and biochemical profiles were studied in mithun bulls and presented in graphical form. Results revealed that FMD vaccination in mithun bulls lead to significantly (p < 0.05) decreased individual motility, live sperm, sperm concentration, acrosomal and plasma membrane integrity in the post–vaccinated mithun bull (Figure 1) whereas total sperm abnormalities were significantly (p < 0.05) increased (Table 1). Antioxidant profiles such GSH, GPD, GRD, CAT, SOD and TAC were decreased whereas total cholesterol and LPO were increased significantly (p<0.05) in semen collected from vaccinated mithun bulls (Figure 2, 3). Similarly the white blood count in blood (Table 1) and rectal temperature (Figure 4) of vaccinated mithun bull were increased as compared to the pre vaccinated mithun bulls.
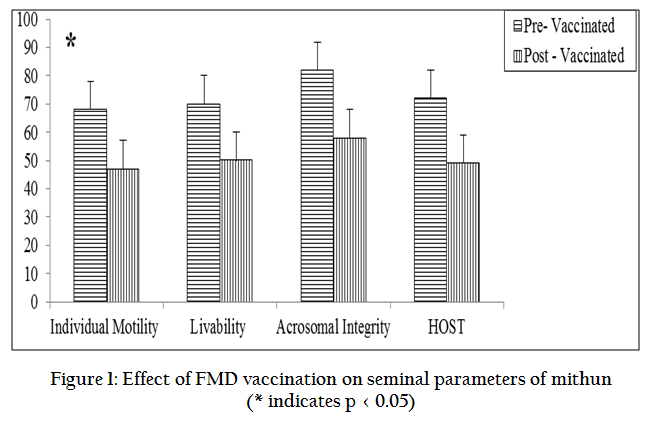

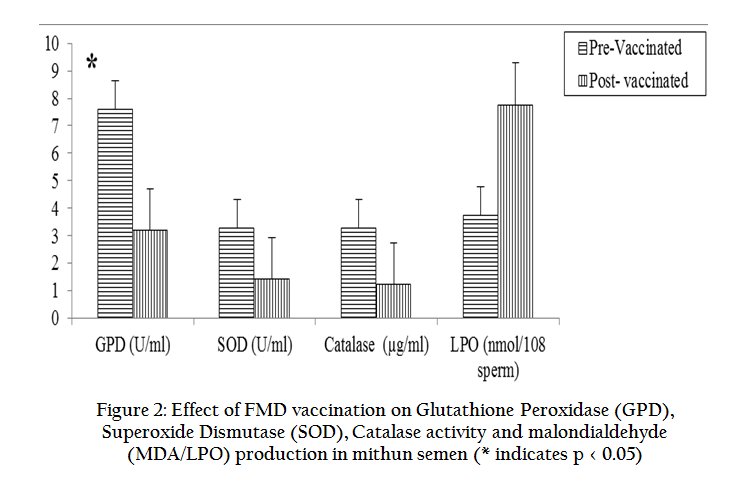
Figure 2: Effect of FMD vaccination on Glutathione Peroxidase (GPD), Superoxide Dismutase (SOD), Catalase activity and malondialdehyde (MDA/LPO) production in mithun semen(* indicates p < 0.05)
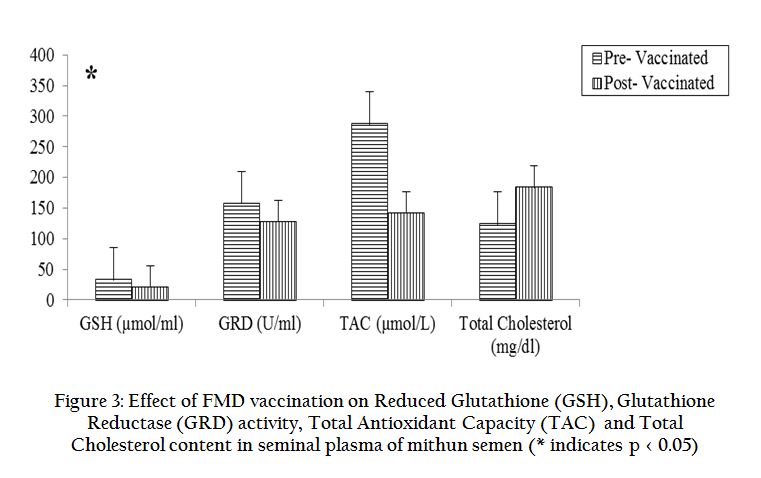
Figure 3: Effect of FMD vaccination on Reduced Glutathione (GSH), Glutathione Reductase (GRD) activity, Total Antioxidant Capacity (TAC) and Total Cholesterol content in seminal plasma of mithun semen (* indicates p < 0.05)
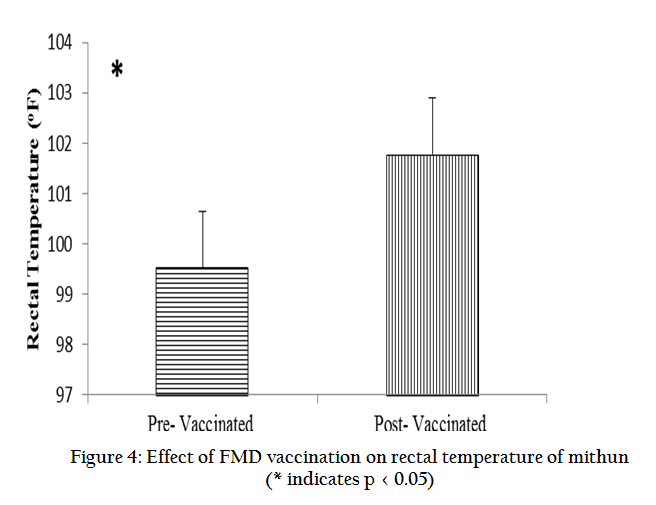
DISCUSSION
In this present study, the results revealed that, vaccination of mithun bulls has affected the seminal parameters, semen quality, antioxidant profiles and biochemical profiles of mithun semen and thus it affects the structures and functional integrity of spermatozoa efficiently. The semen from vaccinated animals is not suitable to preserve for artificial insemination.
Various seminal parameters revealed that volume, colour, mass activity, forward progressive motility, livability, acrosomal integrity and plasma membrane integrity are essential for optimum utilization of semen in frozen semen bank and artificial insemination centre. In this study, FMD vaccination on these seminal parameters revealed significant difference (p< 0.05) between the pre vaccinated and post vaccinated stages. The harmful effects of vaccination in semen preservation are due to it increases the body temperature in general, testes and accessory glands in specifically and crossly it causes seminiferous tubule vacuoles and reduced weight of the testis (Xu et al., 2000).
Sperm individual motility, livability, plasma and acrosomal integrity were decreased and total sperm abnormalities were increased significantly after vaccination was in agreement with previous reports in cattle (Kammar and Gangadhar, 1998; Bhakat et al., 2008; Singh et al., 2003; Bhakat et al., 2010; Venkatareddy et al., 1991) and buffaloes (Pankaj et al., 2007; Bhakat et al., 2010). Motility of spermatozoa develops during their transient passage through the epididymis (Moulikrishnan and Ramamohan Rao, 1986). Anaphylactic effects of FMD vaccination leads to the significant rise in body as well as testes temperature, causes derangement in epididymal functions, storage of sperm and spermatogenesis effectively (Venkatareddy et al., 1991) and that leads to vaccination mediated reduced sperm motility and with similar features of testicular hypoplasia and degeneration (Arthur et al., 1989). Enhanced temperature could lead to raise in the secondary abnormalities (Venkataswami and Rao, 1970) and sperm tail and mid–piece abnormalities as in testicular degeneration or partial hypoplasia of testes (Sullivan, 1978). Low sperm motility is associated with high incidence of sperm tail defects as a result of epididymal dysfunction and poor handling in the laboratory were reported (Rao, 1976).
Harmful effects of vaccination on sperm concentration has been reported in domestic species especially in cattle and buffalo (Bhakat et al., 2008; Venkatareddy et al., 1991; Singh et al., 2003), but it has not been established completely (Pankaj et al., 2007; Kammar and Gangadhar, 1998). The reduced concentration of sperm in vaccinated animal might be due to the harmful effects of therapeutic agents on germinal cells and sperm cell resulting into increase in abnormal and dead spermatozoa, which are phagocytosed subsequently by leucocytes (Mann and Mann, 1981). In mithun bulls, foot and disease vaccination has adverse effects in total motile sperm per ejaculate and day.
The present study reports in the acrosomal integrity were associated with reports of Saxena and Tripathi (1977) and Gowda (1993) and they were reported that intact acrosome percentage was decreased after FMD vaccination. The acrosome in sperm of vaccinated bulls was either detached and/or broken and subsequently release of acrosomal enzymes and leads to reduction of fertilizing capacity of spermatozoa and animal. Vaccination stress induces abnormal acrosomal development as observed in testicular degeneration (Bane and Nicander, 1966).
Plasma membrane integrity (HOST) has significantly (p<0.05) decreased after vaccination (Singh et al., 2004). Because, HOST is an ideal test to detect the biochemical integrity of plasma membrane of sperm and it is also involved in the process of sperm capacitation, acrosome reaction and fertilization of the oocyte (Jayendran et al., 1984). Decreased in percentage of HOST positive sperm was reported in scrotal insulation with thermal stress in bucks due to rise in testicular temperature (Antoine and Pattabiraman, 1999). Similarly, Sivaramalingam (1994) reported that in bulls, HOS test reacting spermatozoa were reduced after heat treatment. Thus it can be concluded that vaccination stress affects the HOST positive sperm percentage by affecting the biochemical structure of the sperm plasma, acrosomal membrane and flagella of spermatozoa. Vaccination thermal stress induces the animal unable to intake of poly unsaturated fatty acids (PUFA) leads to impaired spermatogenesis and membrane transition (Chanmugan et al., 1984) and a overflow in blood circulation of the testis results in local heating, temperature rises to 42oC – 43oC (Mieusset et al., 1992) leads to hypoxic stress to testes and insufficient oxygenation results formation of ROS leads to cell cycle arrest and apoptosis (Iida et al., 2002) and it has been shown to cause depletion of germ cells (Krakowska et al., 2006) and its function (Paul et al., 2009).
Mammalian sperm membrane has high polyunsatured fatty acids, exhibits the sperm very susceptible to ROS by altering of motility and membrane integrity as it changes the phase transition of membrane of sperm and damage to sperm DNA (Griveau et al., 1995; Perumal et al., 2011a; Perumal et al., 2011b) and ultimately affects fertility of mithun bull. It is therefore crucial for the male reproductive system to be well guarded against oxidative injury. Mithun semen normally contains anti–oxidants, including GSH, GRD, GPD, CAT, SOD and TAC that can offset lipid peroxidation (Perumal et al., 2013a; Perumal et al., 2013b). But vaccination induces febrile condition to the whole body and reproductive system. The rate of generation of ROS in testis and/or spermatozoa depends upon temperature of the testis and body (Ikeda et al., 1999) as vaccination leads to thermo stress causes deterioration of sperm quality. Moreover, the concentration of these antioxidants is reduced due to thermal stress and anaphylactic shock in vaccinated animals (Ahotupa and Huhtaniemi, 1992) as these antioxidants are derived from epididymis (Fouchecourt et al., 2000; Zini et al., 2002) especially cauda epididymis (Mueller et al., 1998) and seminal vesicle (Tramer et al., 1998) into the semen as the epididymis, accessory sex glands are thermo sensitive and androgen dependent (Saeed et al., 1994). Several lines of evidence indicate that gross impairment of the endocrinological functions of testes in thermo stress leads to a drastic decrease in the number of LH receptors (Risbridger et al., 1981) and impairment of gonadotropin uptake (Sharpe, 1983) in the testis. Moreover, a decline in the activity of enzymes associated with androgen biosynthesis (Llaurado et al., 1963), and decreased androgen–binding protein production (Kerr et al., 1979) have been noted in this affected animals.
The total white blood cell count was increased in the blood of post vaccinated mithun bulls as because the vaccination has created the thermal and anaphylactic shock / stress which enhanced secretion of adrenal stress hormones (Cortisol), thus the white blood cell count was increased. The vaccination stress has induced changes similar to degeneration or inflammatory condition causes increased seminal lymphocytes and macrophages mainly from the epididymis and rete testis, whereas granulocytes are largely contributed by the prostate and seminal vesicles (El–Demiry et al., 1985) and very rarely granulocytes from secretions of the prostate (Schaeffer et al., 1981). White blood cells in semen can also be an early sign of acute epididymitis (Wolff, 1995) as seen in vaccinated animals. If concentrations of activated granulocytes are elevated in the epididymis, prostate, or seminal vesicles during a silent genital tract infection, the released ROS could impair normal sperm function (Wolff et al., 1991). However, leukocytes are not the only source of ROS, defective spermatozoa with an excessive amount of residual cytoplasm can produce higher amounts of ROS (Ochsendorf, 1999) as we have seen in the semen of vaccinated mithun. Seminal plasma strongly quenches the oxidative bursts released by granulocytes in response to infection or inflammation or degeneration. Mithun with higher antioxidant levels may tolerate larger numbers of ROS–producing WBCs, whereas bulls without adequate seminal plasma protection may suffer sperm damage by granulocyte numbers as low as possible (Kovalski et al., 1992). Moreover effective effect of antioxidant against the ROS will depend on the source and nature of ROS; that is, whether they are produced by abnormal, dead spermatozoa and/or neutrophils in extracellularly or within the spermatozoa in intracellularly.
Glutathione (GSH) is the most prominent non–protein thiol in the mammalian cells and is present mostly in the reduced form (GSH) and only a little amount is present in oxidized form (CSSG). GSH antioxidant system and cycle consists of GSH, GSSG, GRD, GPD, glutathione – s – transferase. Glutathione reductase stimulates the reduction of GSSG to GSH. This will help to steady supply of the reductive substrate (NADPH) to GPD. Glucose –6– phosphate dehydrogenase (G6PD) is intracellular enzymes and required for the conversion of NADP to NADPH, is called GSH oxidizing–reducing cycle in sperm and seminal plasma. It is affected by thermal stress (Ahotupa and Huhtaniemi, 1992) due to vaccination. In this present study GSH, GRD, GPD were reduced in the seminal plasma of mithun as they are positively correlated with sperm parameters suggests that this enzyme is higher in the seminal plasma and low in the spermatozoa of the ejaculated semen (Brown et al., 1977; Perumal et al., 2013b). GRD has catalyses oxidation of GSH to CSSG and thus reduction of H2O2 to H2O in the semen.
Catalase is a tetrameric protein contains four polypeptide chains and is found in all living organisms exposed to aerobic environment condition. This antioxidant is found in the epididymis and seminal vesicle. It detoxifies both the intra and extracellular hydrogen peroxide by conversion of H2O2 to H2O and O2, by eliminating the potential ROS toxicity (Aitken, 1995; Aitken et al., 1994) and it can inhibits the loss of motility caused by leukocyte generated ROS (de Lamirande et al., 1997). In this present study, the concentration of CAT was decreased in vaccinated mithun bulls because the thermo stress affects the normal functions of testes as well as the accessory sex glands as similar in cryptorchid testes (Ahotupa and Huhtaniemi, 1992).
Similarly, SOD dismutates the superoxide into oxygen and hydrogen peroxide in semen. Thus, it acts as an essential antioxidant in mithun semen. It scavenges both extra as well as intracellular superoxide anion and prevents lipid peroxidation of the sperm plasma membrane. SOD dismutase (O2–) spontaneously anion to form O2 and H2O2. SOD prevents premature hyper activation and capacitation induced by ROS before ejaculating (de Lamirande and Gagnon, 1995). In this study the concentration of SOD was decreased in post vaccinated animals as the thermo stress affects the normal functions of testes as well as accessory sex glands (Ahotupa and Huhtaniemi, 1992). These findings of the present study revealed that impaired detoxication of ROS and concomitant oxidative stress effect may be implicated in biochemical mechanisms responsible for testicular dysfunction in vaccinated animals. Retaining the integrity of the highly specialized structure should be of vital importance to sperm function. Excess production of ROS and thermal stress induced by vaccination can be prevented by feeding of sufficient antioxidants in the feed as the supplement (Jayaganthan et al., 2013) and feed or adding of the antioxidants as the additives in the semen preservation process (Perumal et al., 2012a; Perumal et al., 2012b).
Phospholipids and cholesterol are needed to maintain the cell physical integrity as well as ensures fluidity of the cell membrane (Srivastava et al., 2013). Cholesterol plays an important role in the sperm membrane because phospholipids and cholesterol are released from the sperm membrane initiates the key step in the process of capacitation, acrosome reaction and fertilization (Witte and Schafer–Somi, 2007). These changes indicate that cholesterol leaving the sperm membrane and its binding to seminal plasma protein in seminal plasma and female reproductive organs (Therien et al., 1998) and is essential for fertilization and conception. Further, addition of cholesterol to semen diluents prior to defreezing of straws increases sperm resistance to cold stress caused by the freezing–defreezing procedures, preserving sperm motility, viability, acrosomal integrity and fertilization potential of sperm (Moore et al., 2005). In this present study, in vaccinated mithun bulls, total cholesterol concentration was significantly (p< 0.05) higher than in pre–vaccinated bull as the heat stress induces damages and destruction of sperm membrane followed by a rise in dead spermatozoa that leads to a high release of cholesterol, phospholipid and fatty acids molecules in the seminal plasma (Phillip, 1972).
It was concluded from the present study, FMD vaccine has more severe harmful effects on semen quality and antioxidant profiles in mithun bulls. Vaccination results febrile reaction causes alteration in rectal temperature, WBC profiles, seminal parameters and antioxidant profiles as in the testicular degeneration. Alteration of this spermiogram, antioxidants and biochemical profiles following FMD vaccination indicates that in this bovine, the semen collection, processing and preservation should be stopped upto the normal fertility of sperm is restored to avoid the conception failure and repeat breeding from artificial insemination using such semen in artificial insemination and frozen semen bank for mithun.
ACKNOWLEDGEMENTS
The authors are grateful to the Director, National Research Centre on Mithun (ICAR), Jharnapani, Nagaland –797 106, India for providing necessary facilities and supports.
CONFLICT OF INTEREST
The authors have no conflict of interest to declare.
REFERENCES
Ahotupa M and Huhtaniemi I (1992). Impaired detoxification of reactive oxygen and consequent oxidative stress in experimentally cryptorchid rat testis. Biol. Reprod. 46: 1114 –1118.
http://dx.doi.org/10.1095/biolreprod46.6.1114
PMid:1356492
Aitken J (1995). Mechanisms of prevention of lipid peroxidation in human spermatozoa. In: Human acrosome reaction (Eds: Fenichel, P. and Parinaud, J.). pp. 339–353.
Aitken RJ, West K and Buckingham D (1994). Leukocytic infiltration into the human ejaculate and its association with semen quality, oxidative stress, and sperm function. J. Androl. 15: 343–352. PMid:7982803
Antoine D and Pattabiraman SR (1999). Spermatozoa response to hypo–osmotic swelling and freezability tests after testicular degeneration in bucks. Indian J. Anim. Reprod. 20: 54–56.
Arthur GH, Noakes DE and Pearson H (1989). Veterinary Reproduction and Obstetrics. 6th ed., London. Bailliere Tindal, 509–584.
Bane A and Nicander L (1966). Electron and light microscopical studies on spermateliosis in a boar with acrosomal abnormalities. J. Reprod. Fert. 11:133 – 138.
http://dx.doi.org/10.1530/jrf.0.0110133
Barman NN, Sarma DK, Das S and Patgiri GP (1999). Foot–and–mouth disease in wild and semi–domesticated animals of the north–eastern states of India. Indian J. Anim. Sci. 69: 781–783.
Bhakat M, Mohanty TK, Gupta AK, Raina VS, Mondal G and Khan HM (2008). Effect of FMD vaccination on various semen characteristics of Sahiwal bulls. Pak. J. Agri. Sci. 45: 327–332.
Bhakat M, Mohanty TK, Gupta AK, Raina VS, Brahma B, Mahapatra R K and Sarkar M (2010). Effect of FMD vaccination on semen quality parameters in Karan Fries and Murrah buffalo bulls. Tropical Animal Health and Production. 42: 1363–1366.
http://dx.doi.org/10.1007/s11250-010-9593-3
PMid:20509049
Brown DV, Senger PL, Stone SL, Froseth JA and Becker WC (1977). Glutathione peroxidase in bovine semen. J. Reprod. Fert. 50: 117–118.
http://dx.doi.org/10.1530/jrf.0.0500117
PMid:864635
Buckett WM, Luckas MJ, Aird IA, Farquharson RG, Kingsland CR and Lewis–Jones DI (1997). The hypo–osmotic swelling test in recurrent miscarriage. Fertil. Steril. 68: 506 –509.
http://dx.doi.org/10.1016/S0015-0282(97)00241-0
Buege JA and Aust SD (1978). Microsomal lipid peroxidation. Methods Enzymol. 52: 302– 310.
http://dx.doi.org/10.1016/S0076-6879(78)52032-6
Chanmugan P, Wheeler C and Hwang DH (1984). Fatty acid composition of the testes of zinc–deficient rats: the effect of docosapentaenoic acid supplementation. J. Nutr. 114: 2073 –2079.
De Lamirande E and Gagnon C (1995). Impact of reactive oxygen species on spermatozoa: A balancing act between beneficial and detrimental effects. Human Reprod. 10: 15–21.
http://dx.doi.org/10.1093/humrep/10.suppl_1.15
PMid:8592032
de Lamirande E, Leclerc P, Gagnon C (1997). Capacitation as a regulatory event that primes spermatozoa for the acrosome reaction and fertilization. Mol. Hum. Reprod. 3:175 – 194.
http://dx.doi.org/10.1093/molehr/3.3.175
PMid:9237244
Dutta PK, Mahanta PN and Chakrabarty AK (1979). Foot–and–mouth disease in mithun (Bos gaurus). Vet. Rec. 104:146.
http://dx.doi.org/10.1136/vr.104.7.146
PMid:452352
El–Demiry MIM, Hargreave TB, Busuttil A, James K, Ritchie AW and Chisolm GD (1985). Lymphocyte sub–populations in the male genital tract. Br. J. Urol. 57: 769 –774.
http://dx.doi.org/10.1111/j.1464-410X.1985.tb07051.x
PMid:3910154
Fouchecourt S, Metayer S, Locatelli A, Dacheux F and Dacheux JL (2000). Stallion epididymal fluid proteome: qualitative and quantitative characterization; secretion and dynamic changes of major proteins. Biol. Reprod. 62: 1790–1803.
http://dx.doi.org/10.1095/biolreprod62.6.1790
PMid:10819784
Gowda BK (1993). Effect of foot and mouth disease vaccination on pre– and post–freezing semen parameters. M.V.Sc. Thesis, University of Agricultural Science, Bangalore, India.
Griveau JF, Dumont E, Renard P, Callegari JP and Le Lannou D (1995). Reactive oxygen species, lipid peroxidation and enzymatic defence systems in human spermatozoa. J. Reprod. Fertil. 103: 17–26.
http://dx.doi.org/10.1530/jrf.0.1030017
PMid:7707295
Hegde R, Gomes AR, Giridhar P, Venkatesh MD and Renukaprasad C (2011). A note on FMD in Mithun (Bos frontalis) and Black buck (Antelope cervicapra). Zoo's Print. 26(6): 19.
Iida T, Mine S, Fujimoto H, Suzuki K, Minami Y and Tanaka Y (2002). Hypoxia–inducible factor–1alpha induces cell cycle arrest of endothelial cells. Genes. Cells. 7: 143–149.
http://dx.doi.org/10.1046/j.1356-9597.2001.00512.x
PMid:11895478
Ikeda M, Kodama H, Fukuda J, Shimizu Y, Murata M, Kumagai J and Tanaka T (1999). Role of radical oxygen species in rat testicular germ cell apoptosis induced by heat stress. Biol. Reprod. 61: 393–399.
http://dx.doi.org/10.1095/biolreprod61.2.393
PMid:10411517
Jayaganthan P, Perumal P, Balamurugan TC, Verma RP, Singh LP, Pattanaik AK and Kataria M (2013). Effects of Tinospora cordifolia supplementation on semen quality and hormonal profile of ram. Animal Reproduction Science 140 (1): 47–53.
http://dx.doi.org/10.1016/j.anireprosci.2013.05.003
PMid:23755935
Jeyendran RS, Vander Ven HH, Perez–Pelaz M, Crabo BG and Zanefeld LJD (1984). Development of an assay to assess the functional integrity of the human sperm membrane and its relationship to other semen characteristics. J. Repod. Fertil. 70: 219–228.
http://dx.doi.org/10.1530/jrf.0.0700219
PMid:6694140
Kammar NF and Gangadhar KS (1998). Effect of foot–and–mouth vaccination on reaction time and some seminal characteristics in Surti bulls. Indian J. Anim. Reprod. 19:149– 150.
Karunakaran M, Dhali A, Mech A, Khate K, Rajkhowa C and Mishra DP (2007). Preservation of mithun (Bos frontalis) semen at refrigeration temperature. Animal Reproduction Science. 101:257– 264
http://dx.doi.org/10.1016/j.anireprosci.2006.09.011
PMid:17095169
Kerr JB, Rich KA and de Kretser DM (1979). Alterations of the fine structure and androgen secretion of the interstitial cells in the experimentally cryptorchid rat testis. Biol. Reprod. 20: 409 – 422.
http://dx.doi.org/10.1095/biolreprod20.3.409
PMid:36931
Kovalski N, de Lamirande E and Gagnon C (1992). Reactive oxygen species generated by human neutrophil inhibit sperm motility: protective effect of seminal plasma and scavengers. Fertil. Steril. 58: 809 – 816.
PMid:1426329
Krakowska M, Bomba G, Jakubowski K, Rotkiewicz T, Jana B and Penkowski A (2006). Impact of oxidative stress and supplementation with vitamins E and C on testes morphology in rats. J. Reprod. Dev. 52: 203–209.
http://dx.doi.org/10.1262/jrd.17028
Llaurado IG and Dominiquez OV (1963). Effect of cryptorchidism on testicular enzymes involved in androgen biosynthesis. Endocrinology. 72: 292–295.
http://dx.doi.org/10.1210/endo-72-2-292
PMid:13931095
Mangurkar BR, Phadnis YP and Bhosrekar MR (2000). Effect of Foot and mouth disease vaccination on semen characteristics of exotic and cross–bred bulls. Indian J. Anim. Reprod. 21: 135–137.
Mann T and Mann CL (1981) Male Reproductive Function and Semen. Springer – Verlag Berlin. Heidelberg, New York, USA.
Mieusset R, Quintana–Casares P, Sanchez–Partida LG, Sowerbutts SF, Zupp JL and Setchell BP (1992). Effects of heating the testes and epididymides of rams by scrotal insulation on fertility and embryonic mortality in ewes inseminated with frozen semen. J. Reprod. Fertil. 94: 337–343.
http://dx.doi.org/10.1530/jrf.0.0940337
PMid:1593536
Mondal SK and Pal DT (1999). Mithun: historical perspective. Asian Agri–Hist. 3: 245–260.
Moore AI, Squires EL and Graham JK (2005). Adding cholesterol to the stallion sperm plasma membrane improves cryosurvival. Cryobiology. 51: 241–249.
http://dx.doi.org/10.1016/j.cryobiol.2005.07.004
PMid:16122725
Moulikrishnan K and Ramamohan Rao A (1986). Epididymal dysfunction in buffalo bulls. Indian Vet. J. 63: 1013–1016.
Mueller A, Hermo L and Robaire B (1998). The effects of aging on the expression of glutathione S–transferases in the testis and epididymis of the Brown Norway rat. J. Androl. 19:450 – 465.
PMid:9733148
Murugavel K, Veerapandian C and Subramanian A (1997). Effect of black quarter vaccination on semen quality in Murrah bulls. Indian J. Anim. Sci. 67: 597 –598.
Ochsendorf FR (1999). Infections in the male genital tract and reactive oxygen species. Hum. Reprod. Update. 5: 399–420.
http://dx.doi.org/10.1093/humupd/5.5.399
PMid:10582780
OIE/FAO FMD Reference Laboratory (2010). Serotype O virus lineages circulating in India during 2008–2010. Annual Report– 2010, 23.
Pankaj PK, Raina VS, Roy B, Mohanty TK and Mishra A (2007). Studies on effect of vaccination on seminal characteristics of riverine buffaloes (Bubalus bubalis). Buffalo Bulletin. 26: 91– 103.
Paul C, Teng S and Saunders PT (2009). A single, mild, transient scrotal heat stress causes hypoxia and oxidative stress in mouse testes, which induces germ cell death. Biol. Reprod. 80: 913–919.
http://dx.doi.org/10.1095/biolreprod.108.071779
PMid:19144962 PMCid:PMC2709966
Perumal P, Selvaraju S, Selvakumar S, Barik AK, Mohanty DN, Das S, Das RK and Mishra PC (2011a). Effect of pre–freeze addition of cysteine hydrochloride and reduced glutathione in semen of crossbred jersey bulls on sperm parameters and conception rates. Reprod. Domestic Anim. 46: 636–641.
http://dx.doi.org/10.1111/j.1439-0531.2010.01719.x
PMid:21134005
Perumal P, Selvaraju S, Barik AK, Mohanty DN, Das S and Mishra PC (2011b). Role of reduced glutathione in improving post–thawed frozen seminal characters of poor freezable Jersey crossbred bull semen. Indian J. Anim. Sci. 81: 807–810.
Perumal P, Selvaraju S, Barik AK, Mohanty DN, Das S, Mishra PC and Veeraselvam M (2012a). Cysteine hydrochloride on post–thawed seminal characters of poor freezable Jersey crossbred bull semen. International Journal of Bio – Resource and Stress Management. 3(2): 232 –235.
Perumal P, Selvaraju S, Barik AK, Mohanty DN, Das S, Mishra PC and Veeraselvam M (2012b). Reduced glutathione and cysteine hydrochloride on sperm motility and velocity parameters of poor crossbred bull semen. International Journal of Bio – Resource and Stress Management. 3(2): 145 –151.
Perumal P, Vupru K and Rajkhowa C (2013a). Effect of addition of taurine on the liquid storage (5oc) of mithun (Bos frontalis) semen. Veterinary Medicine International 2013: 7 pages, Article ID 165348.
Perumal P, Vupru K and Rajkhowa C (2013b). Effect of reduced glutathione on the liquid storage (5oc) of mithun (Bos frontalis) semen. Indian J. Anim. Sci. 83: 1024 – 1028.
Phillip MC (1972). The physical state of phospholipids and cholesterol in monolayers, bilayers and membrane. Prag. Surface Membr. 5: 139– 142.
http://dx.doi.org/10.1016/B978-0-12-571805-9.50009-9
Rajkhowa S, Rajkhowa C and Bujarbaruah KM (2003). Diseases of mithun (Bos frontalis) a revised. Veterinary Bulletin. 73: IR–6R.
Rao AR (1976). Sperm akinesia in a hollikar bull. Indian Vet. J. 53; 414 – 418.
PMid:1026657
Risbridger GP, Kerr JB, Peake R, Rich KA and de Kretser DM (1981). Temporal changes in rat Leydig cell function after the induction of bilateral cryptorchidism. J. Reprod. Fertil. 63:415 – 423.
http://dx.doi.org/10.1530/jrf.0.0630415
PMid:6117660
Robeck TR and O'Brien JK (2004). Effect of cryopreservation methods and precryopreservation storage on bottlenose dolphin (Tursiops truncatus) sperms. Biol. Reprod. 70: 1340–1348.
http://dx.doi.org/10.1095/biolreprod.103.025304
PMid:14711792
Saeed S, Khan FA, Rehman SB, Khan DA and Ahmad M (1994). Biochemical parameters in evaluation of oligospermia. J. Pak. Med. Assoc. 44:137–140.
PMid:8089906
Saxena VB and Tripathi SS (1977). Effect of foot and mouth disease vaccination on semen quantity, quality and preservability in Jersey bulls. Indian Vet. J. 54: 959 – 964.
Schaeffer AJ, Wendel EF, Dunn JK and Grayhack JT (1981). Prevalence and significance of prostatic inflammation. J. Urol. 125: 215–220.
PMid:7206060
Sharpe RM (1983). Impaired gonadotrophin uptake in vivo by the cryptorchid rat testis. J. Reprod. Fertil. 67: 379–387.
http://dx.doi.org/10.1530/jrf.0.0670379
PMid:6132004
Simoons FJ (1984). Gayal or mithun. In: Manson, I.L. (Ed.), Evolution of Domesticated Animals. Longman, London, 34–36.
Singh R, Verma HK and Kumar S (2003). Effect of the foot and mouth disease vaccination on the semen quality of buffalo bulls. Indian J. Anim. Sci. 73: 1319–1323.
Singh R, Verma HK, Kumar S and Matharoo JS (2004). Effect of foot and mouth disease vaccination on the reproductive capacity of the buffalo bulls. Indian J. Anim. Reprod. 25; 45 – 47.
Sivaramalingam K (1994). Studies on hypo–osmotic swelling test (HOST) for evaluation of bull spermatozoa. M.V.Sc. Thesis, Madras Veterinary College, Madras. India.
Srivastava N, Srivastava SK, Ghosh SK, Amit Kumar, Perumal P and Jerome A (2013). Acrosome membrane integrity and cryocapacitation are related to cholesterol content of bull spermatozoa. Asian Pacific Journal of Reproduction 2(2): 126–131.
http://dx.doi.org/10.1016/S2305-0500(13)60132-3
Suleiman SA, Ali ME, Zaki MS, Malik EME and Nast MA (1996). Lipid peroxidation and human sperm motility: protective role of vitamin E. J. Androl. 17: 530–537.
PMid:8957697
Sullivan JJ (1978). Morphology and motility of spermatozoa. In: Salisbury, G.W., Van Demark, N.L. and Lodge, J.R. (ed): Physiology of Reproduction and Artificial Insemination of Cattle. (2nd Edn). San Francisco, W.H. Free Man, 286–328.
Therien I, Moreau R and Manjunath P (1998). Major proteins of bovine seminal plasma and high density lipoproteins induce cholesterol efflux from epididymal sperm. Biol. Reprod. 59: 768–776.
http://dx.doi.org/10.1095/biolreprod59.4.768
PMid:9746724
Tomar NS (1997). Artificial Insemination and Reproduction of Cattle and Buffalos. Sarojprakashan, Allahabad, India.
Tramer F, Rocco F, Micali F, Sandri G and Panfili E (1998). Antioxidant systems in rat epididymal spermatozoa. Biol. Reprod. 59: 753–758.
http://dx.doi.org/10.1095/biolreprod59.4.753
PMid:9746722
Venkatareddy J, Venkatamunichetty A, Ramachandran SV and Sreeraman V (1991). Effect of foot and mouth disease vaccination on semen quality. Indian J. Anim. Reprod. 12: 13–14.
Venkataswami V and Rao VJ (1970). Preliminary report on the effect of foot and mouth disease vaccination on semen quality of crossbred bulls. Indian Vet. J. 47: 23–29.
PMid:4314816
Verma ND and Sarma DK (1997). Note on foot–and–mouth disease in mithun in Arunachal Pradesh. Indian J. Virol. 13: 75–76.
Watson PF (1975). Use of Giemsa Stain to detect change in acrosome of frozen ram spermatozoa. Veterinary Record. 97: 12–15.
http://dx.doi.org/10.1136/vr.97.1.12
PMid:49974
Witte TS and Schafer–Somi S (2007). Involvement of cholesterol, calcium and progesterone in the induction of capacitation and acrosome reaction of mammalian spermatozoa. Anim. Reprod. Sci. 102: 181–193.
http://dx.doi.org/10.1016/j.anireprosci.2007.07.007
PMid:17870257
Wolff H (1995). The biologic significance of white blood cells in semen. Fertil. Steril. 63: 1143–1157.
PMid:7750580
Wolff H, Bezold G, Zebhauser M and Meurer M (1991). Impact of clinically silent inflammation on male genital tract organs as reflected by biochemical markers in semen. J. Androl. 12: 331–334.
PMid:1765569
Xu L, Sun H, Wang S, Song L, Chang HC and Wang X (2005). The roles of metallothionein on cadmium–induced testes damages in Sprague–Dawley rats. Environ. Toxicol. Pharmacol. 20: 83–87.
http://dx.doi.org/10.1016/j.etap.2004.10.008
PMid:21783572
Zini A, Fischer MA, Mak V, Phang D and Jarvi K (2002). Catalase–like and SOD–like activities in human seminal plasma. Urol. Res. 30: 321–323.
http://dx.doi.org/10.1007/s00240-002-0283-0
PMid:12389121

