
Advances in Animal and Veterinary Sciences


Short Communication
Advances in Animal and Veterinary Sciences 1 (1): 41–43Sequence and Phylogenetic Analysis of Bovine Rotavirus Isolates (G6 Genotypes) from India
Yashpal S. Malik*, Naveen Kumar, Kuldeep Sharma, AA Haq, Amit Kumar, Minakshi Prasad
-
Division of Biological Standardization, Indian Veterinary Research Institute, Izatnagar, Uttar Pradesh, India
*Corresponding author:[email protected]; [email protected]
ARTICLE CITATION:
Malik YS, Kumar N, Sharma K, Haq AA, Kumar A, Prasad M (2013). Sequence and phylogenetic analysis of bovine rotavirus isolates (G6 genotypes) from India. Adv. Anim. Vet. Sci. 1 (1): 41–43
Received: 2013–04–23, Revised: 2013–04–27, Accepted: 2013–04–28
The electronic version of this article is the complete one and can be found online at
(
http://www.nexusacademicpublishers.com/table_contents_detail/4/35/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Rotaviruses (RVs) are among the most important enteric pathogens responsible for significant morbidity and mortality in bovine calves throughout the world. We describe here the sequence characterization of bovine group A rotavirus isolate BR108 obtained from Northern India (semi–arid zone) and P–43 isolate isolated from foot–hills of western Himalayan region (Uttarakhand). The isolates were characterized by phylogenetic analysis of one of its major outer capsid protein gene, VP7. Analysis of VP7 gene sequences of both the isolates (BR–108 and P–43) revealed higher sequence homology of 95.6% at nucleotide and 98.3% at amino acid level with bovine rotavirus G6 genotype from Punjab. The findings are suggestive of inter–geographical boundaries of states transmission of bovine rotaviruses which along with diverse genotypes put a heavy burden on bovine population.
The foremost cause of diarrhoeal diseases in animals worldwide is mainly attributed by group A rotaviruses causing huge economic losses especially in developing countries (Estes and Kapikian, 2007). Rotaviruses (RVs) having genome of 11 segments of double stranded RNA encodes six structural and six non–structural proteins. Two structural proteins (VP4 and VP7) bearing major antigenic determinants have been used widely for the classification of RVs in genotypes, dividing VP7 into 27 G genotypes and VP4 into 35 P genotypes (Matthijnssens et al., 2011). Recently, the remaining rotavirus genes encoding VP1, VP2, VP3, VP6, NSP1, NSP2, NSP3, NSP4 and NSP5 have also been divided into various genotypes, based on specific nucleotide sequence cut–off identity values for each gene segment (Maes et al., 2009). In India, G6, G8 and G10 genotypes of bovine group A rotaviruses are epidemiological important (Gulati et al., 1999, Wani et al., 2004, Bhat et al., 2005, Saravanan et al., 2006, Manuja et al., 2008, Dhama et al., 2009, Minakshi et al., 2009, Singh and Jhala, 2011) but G3 genotype has emerged dominantly in bovine population since last decade (Varshney et al., 2002, Ghosh et al., 2007, Malik et al., 2012).
Characterization of VP7 gene of bovine group A rotaviruses is important to the point of genotypic prevalence, interspecies transmission and formulation of vaccine. Literature search revealed that sequence analysis of VP7 gene of rotavirus is still in infancy in India, as only a few authors have analyzed the VP7 gene sequences of bovine rotaviruses (Varshney et al., 2002; Ghosh et al., 2008; Manuja et al., 2008; Malik et al., 2012). Sequence analysis of VP7 gene gives an insight into the possible origin/relatedness with the rotaviruses of different geographical locations or species. Keeping the importance of VP7 gene sequence analysis, the present study was undertaken to characterize the VP7 gene of two bovine rotavirus isolates collected from different geographical locations i.e. semi–arid area (Haryana) and plains of temperate western Himalayan area (Uttarakhand).
The diarrhoeic faecal samples, one each from Haryana (BR–108) and Uttarakhand (P–43) were retrieved from Enteric Virus Laboratory, IVRI, Mukteswar and 10% faecal suspension (w/v) was prepared in phosphate buffered saline (0.01 M, pH 7.4; Sigma, USA). To remove the coarse particulate matter samples were centrifuged at 2000 g for 10 min and the upper aqueous layer was filtered through a 0.22 μm syringe filter (MDI, India). The suspension was archived and stored at –20oC until further use.
The viral RNA was extracted from 500 μl faecal suspension using an equal volume of TriReagent–LS (Sigma, St. Louis, USA) following manufacturer’s instructions. The extracted RNA was assessed qualitatively and quantitatively using a Nanodrop Spectrophotometer (ND–1000, ThermoScientific, USA) and stored at –20oC until further use.
The extracted viral dsRNA was analyzed by PAGE which was performed by using 5% stacking and 10% resolving gels as per the method described in our previous study (Malik et al., 2012). Briefly, extracted RNA (200–500 ng) was mixed with equal amount of 2X RNA loading buffer and loaded in the wells. The gel was electrophorosed at 120V in 1X Tris–Glycine buffer. Subsequently, the gel was impregnated with silver nitrate following the method described previously (Malik et al., 2012).
The cDNA was synthesized from 5 µl extracted RNA using 20U of M–MuLV, 10mM of each dNTPs, 0.2 µg Random Hexamer primer, 2 µl Dimethyl sulphoxide (DMSO) at 37ºC for one hour and final denaturation at 80ºC for 3 min. The first round of PCR was carried out for the amplification of full length (P–43) and partial length (BR–108) of VP7 gene as described in our previous study (Malik et al., 2012). The second round PCR was done using a cocktail of primers for the genotypes viz. G3, G6, G8 and G10 using 1:100 dilutions of first round PCR amplicons as template. Second round amplicons of multiplex nested PCR were resolved by agarose gel electrophoresis (2% w/v) at 100 V for 1 hour in 1X TAE buffer with 0.5 µg/ml ethidium bromide and viewed under Gel Documentation system–ImageMaster® VDS (Syngene, DSS, UK).
The second round multiplex nested PCR amplicons were gel–purified using GeneJet Gel Extraction Kit (Fermentas, EU). The gel–purified PCR amplicons were cloned into the pGEM–T Easy Vector cloning system (Promega) and sequenced by SciGenom Labs Pvt. Ltd., Kerala. Phylogenetic analysis was performed using MEGA 5.05 software. Briefly, the nucleotide sequences of VP7 gene of both the samples were aligned by Clustal W with the available VP7 gene sequences in the NCBI database of animal RVA from India and rest of the world. The percent identity and divergence of the nucleotide and deduced amino acid sequence was performed to determine the percentage homology with other RVA VP7 gene sequences from different geographical locations. Phylogenetic tree was constructed by neighbor–joining statistical method. Rota C v2.0 web based tool for rotavirus classification (http://rotac.regatools.be) was also used for the confirmation of the sequence obtained and automated genotyping based on VP7 sequences.
The sequences of VP7 gene of the rotavirus BR–108 and P–43 strain characterized here have been submitted in the GenBank database and assigned accession numbers, JF689842 and HM591496, respectively.
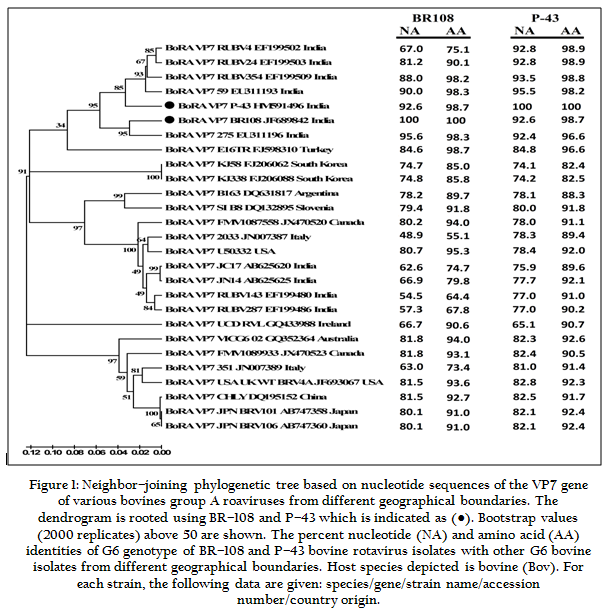
Figure: Neighbor–joining phylogenetic tree based on nucleotide sequences of the VP7 gene of various bovines group
The RNA–PAGE analysis of both the bovine rotavirus isolates revealed long electropherotypes with 11 dsRNA segments migrating in a pattern of 4:2:3:2, where segments 7, 8, 9 moved in a triplet, typical of bovine group A rotaviruses. Further, genomapping for G3, G6, G8 and G10 genotypes was done for both these positive samples by multiplex nested PCR. The second round of multiplex nested PCR of both the positive samples showed 581 bp specific amplicons on resolving on 2% agarose gel revealing the confirmation of isolates being of G6 genotype. For the VP7 sequences comparison, VP7 sequences of both the group A bovine rotavirus isolates, one each from Haryana and Uttarakhand states were aligned with the retrieved VP7 sequences from NCBI database originating from the rotaviruses of different geographical locations of the world. The sequence alignment by clustal W and phylogenetic analysis by Neighbour–joining method was performed with the MEGA 5.05 software. The VP7 sequence homology analysis of BR–108 isolate of Haryana revealed maximum homology (95.6 % at nucleotide level and 98.3% at amino acid level) with the bovine group A rotavirus isolate of Punjab (Accession number: EU311196) as shown in Fig.1. Similarly, VP7 sequence homology analysis of P–43 isolate of Uttarakhand revealed maximum homology (95.5 % at nucleotide level and 98.2% at amino acid level) with the bovine group A rotavirus isolate of Punjab (Accession number: EU311193). Analysis of both the bovine rotavirus isolates namely BR–108 and P–43 clustered with bovine rotavirus of Punjab.
Genotyping by multiplex nested PCR is a well–established and recognized epidemiological tool for examining rotavirus strain diversity (Desselberger et al., 2001). In recent years, it has been shown to be the most useful assay for the genotyping of rotavirus of different species, but genotyping without phylogenetic analysis provides information only about the prevalence of different rotavirus genotypes. DNA sequencing followed by sequence comparison and phylogenetic analysis add the vital information about the possible origin of particular gene of rotavirus from different geographical locations or species which is valuable in understanding the evolution of rotaviruses. Therefore, the present study was undertaken to understand the possible origin of two bovine rotaviruses isolates namely BR–108 (Haryana) and P–43 (Uttarakhand). Both the bovine rotavirus isolates in RNA–PAGE analysis revealed long electropherotypes based on migration of segment 10 and 11. The pattern was of RNA segments migration of 4:2:3:2 was typical of group A rotavirus type. Genotyping following our previous optimized protocols for G3, G6, G8 and G10 genotypes by multiplex nested PCR showed 581 bp specific amplicons representing G6 genotype. It is one of the most important genotype epidemiologically after G10 genotype (Gulati et al., 1999, Wani et al., 2004, Bhat et al., 2005, Saravanan et al., 2006, Manuja et al., 2008, Minakshi et al., 2009, Singh and Jhala, 2011) indicating its significant role in provoking havoc in newborn calves by causing fatal diarrhoea. The G6 genotypes have been reported both as single G6 genotype at a rate of 5.56%–26.67% (Gulati et al., 1999; Minakshi et al., 2005; Manuja et al., 2008) and in combination with G10 (Gulati et al., 1999; Minakshi et al., 2009; Sharma et al., 2009) at a rate of 8.3%–11.1% from the Northern part of India. The genotyping as well as sequence analysis of VP7 gene of bovine group A rotavirus is critical not only in vaccine formulation but also to identify its possible origin. Only a limited authors have made an attempt to compare VP7 sequence of bovine group A rotavirus with other bovine rotavirus G3 genotype (Malik et al., 2012), G10 genotype (Varshney et al., 2002), G15 genotype (Ghosh et al., 2008), G6 and G10 (Manuja et al., 2008).
As shown in Fig.1, the VP7 sequence analysis of both the group A bovine rotavirus isolates revealed maximum homology (95.6 % at nucleotide level and 98.3% at amino acid level) with the bovine group A rotavirus isolate of Punjab (Accession number: EU311196). Similarly, VP7 sequence homology analysis of P–43 isolate of Uttarakhand revealed maximum homology (95.5 % at nucleotide level and 98.2% at amino acid level) with the bovine group A rotavirus isolate of Punjab (Accession number: EU311193). Analysis of both the bovine rotavirus isolates namely BR–108 and P–43 clustered with bovine rotavirus of Punjab. The close homology of VP7 gene of both the bovine rotavirus isolates with the bovine rotavirus isolate of Punjab suggests the frequent migration of infected animals across the geographical boundaries. Geographically, Haryana share a large portion of its border with Punjab and Uttarakhand share its border with Haryana which allow easy access for the migration of infected animals to Haryana and Uttarakhand states from Punjab. This favors the rotavirus to proliferate its kingdom over bovines by crossing the geographical boundaries and various species creating an urgent need of intensive surveillance programme to know the exact prevalence of genotypes circulation in bovine population and also limiting the migration of infected animals across the geographical boundaries so as to contain the burden of rotaviruses in bovine population.
ACKNOWLEDGEMENTS
The authors acknowledge the financial help in the form of a project from Uttarakhand State Biotechnology Department, Dehradun and the Director, Indian Veterinary Research Institute for the infrastructural help.
REFERENCES
Bhat MA, Wani SA, Taku A and Dutta TK (2005). Molecular epidemiology of rotavirus infection in animals in India: An update. Indian Journal of Animal Sciences, 75(11): 1276-1282.
Desselberger U, Gray J and Estes MK (2005). Rotaviruses. In: B.W.J. Mahy and V.T. Meulen (eds), Topley and Wilson's Microbiology and Microbial infections, ASM press, USA, 946–958.
Dhama K, Chauhan RS, Mahendran M and Malik SVS (2009). Rotavirus diarrhea in bovines and other domestic animals. Veterinary Research Communication, 33: 1–23.
http://dx.doi.org/10.1007/s11259-008-9070-x
PMid:18622713
Estes M and Kapikian A (2007). Rotaviruses. In: Knipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE, editors. Fields virology. Philadelphia: Kluwer Health/Lippincott, Williams and Wilkins. p. 1917–1974.
Ghosh S, Samajdar S, Sinha M, Kobayashi N, Taniguchi K and Naik TN (2008). Molecular characterization of rare bovine group A rotavirus G15P[11] and G15P[21] strains from eastern India: identification of simian SA11-like VP6 genes in G15P[21] strains. Virus Genes, 37: 241–249.
http://dx.doi.org/10.1007/s11262-008-0260-y
PMid:18679786
Ghosh S, Varghese V, Samajdar S, Sinha M, Kobayashi N and Naik TN (2007). Molecular characterization of bovine group A rotavirus G3P[3] strains. Archives of Virology, 152: 1935-1940.
http://dx.doi.org/10.1007/s00705-007-1009-y
PMid:17577612
Gulati BR, Nakagomi O, Koshimura Y, Nakagomi T and Pandey R (1999). Relative frequencies of g and p types among rotaviruses from Indian diarrheic cow and buffalo calves. Journal of Clinical Microbiology, 37(6): 2074–2076.
PMid:10325385 PMCid:PMC85038
Maes P, Matthijnssens J, Rahman M and Ranst MV (2009). RotaC: A web-based tool for the complete genome classification of group A rotaviruses. BMC Microbiology, 9(238): 1-4.
Malik YPS, Sharma K, Vaid N, Chakravarti S, Chandrashekar KM, Basera SS, Singh R, Minakshi, Prasad S, Gulati BR, Bhilegaonkar KN and Pandey AB (2012). Frequency of group A rotavirus with mixed G and P genotypes in bovines: predominance of G3 genotype and its emergence in combination with G8/G10 types. Journal of Veterinary Science, 13(3): 271-278.
http://dx.doi.org/10.4142/jvs.2012.13.3.271
PMid:23006956 PMCid:PMC3467402
Manuja BK, Prasad M, Manuja A, Gulati BR and Prasad G (2008). A novel genomic constellation (G10P[3]) of group A rotavirus detected from buffalo calves in northern India. Virus Research, 138: 36–42.
http://dx.doi.org/10.1016/j.virusres.2008.08.006
PMid:18793686
Matthijnssens J, Ciarlet M, McDonald SM, Attoui H, Banyai K, Brister JR, Buesa J, Esona MD, Estes MK, Gentsh JR, Iturriza-Gomara M, Johne R, Kirkwood CD, Martella V, Mertens PPC, Minakshi, Prasad G and Grover YP (2009). Occurrence of dual infection of bovine group A rotavirus in diarrhoeic calf in Haryana, India. Indian Journal of Animal Sciences, 79 (12): 1205–1208.
Minakshi, Prasad G, Malik Y and Pandey R (2005). G and P genotyping of bovine group A rotaviruses in faecal samples of diarrhoeic calves by DIG-labeled probes. Indian Journal of Biotechnolgy, 4: 93-99.
Nakagomi O, Parreno V, Rahman M, Ruggeri FM, Saif LJ, Santos N, Steyer A, Taniguchi K, Patton JT, Desselberger U and Van Ranst M (2011). Uniformity of rotavirus strain nomenclature proposed by the Rotavirus Classification Working Group (RCWG). Archives of Virology, 156(8): 1397-1413.
http://dx.doi.org/10.1007/s00705-011-1006-z
PMid:21597953 PMCid:PMC3398998
Saravanan M, Parthiban M and Ramdass P (2006). Genotyping of rotavirus of neonatal calves by nested-multiplex PCR in India. Veterinarski Arhiv, 76: 497–505.
Sharma G, Taku A, Chhabra R and Bhat MA (2009). Prevalence of bovine rotaviral diarrhoea in Jammu. Indian Journal of Virology, 20: 53-58.
Singh TC and Jhala MK (2011). G-typing of bovine rotaviruses by using VP7 gene specific heminested RT-PCR from diarrhoeic calf faecal samples. Buffalo Bulletin, 30(2): 113-138.
Varshney B, Jagannath MR, Vethanayagam RR, Kodhandharaman S, Jagannath HV, Gowda K, Singh DK and Rao CD (2002). Prevalence of, and antigenic variation in, serotype G10 rotaviruses and detection of serotype G3 strains in diarrheic calves: implications for the origin of G10P11 or P11 type reassortant asymptomatic strains in newborn children in India. Archives of Virology, 147: 143-165.
http://dx.doi.org/10.1007/s705-002-8308-z
PMid:11855628
Wani SA, Bhat MA, Ishaq SM and Ashrafi MA (2004). Determination of bovine rotavirus G genotypes in Kashmir, India. Revue Scientifique Et Technique De L`Office International Des Epizooties, 23(3): 931-936.

