
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 2 (7): 377 – 380Analysis of immune response genes in Peripheral Blood Mononuclear Cells (PBMCs) of commercial and indigenous chicken breeds
Arun Saravana Kumar Annamalai, Swati Sachan, Saravanan Ramakrishnan*, Bal Krishan Sharma, Anand Kumar, Vimal Kumar, Surendra Kumar Badasara
-
Immunology Section, Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh 243122
*Corresponding author:[email protected]
ARTICLE CITATION:
Annamalai ASK, Sachan S, Ramakrishnan S, Sharma BK, Kumar A, Kumar V, Badasara SK (2014). Analysis of immune response genes in Peripheral Blood Mononuclear Cells (PBMCs) of commercial and indigenous chicken breeds. Adv. Anim. Vet. Sci. 2 (7): 377 – 380.
Received: 2013–07–01, Revised: 2014–07–18, Accepted: 2014–07–19
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/journal.aavs/2014/2.7.377.380
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Cytokines, a family of small proteins secreted by cells play a major and decisive role in immunity and has pleiotropic effects on different cells. Peripheral Blood Mononuclear Cells (PBMCs) are important cells which play critical role in activation of both innate and adaptive immune responses. Cytokine expression varies with different breeds of chickens and this expression plays a critical role in modulation of immune responses in host. In our study, the basal level expression of different cytokines were assessed in chicken PBMCs (ex vivo) by quantitative real time PCR and comparison between different breeds were determined namely broiler, White Leghorn, and Indigenous Kadaknath. Type I Interferon (IFN) (IFN-β), Th1 cytokine (IFN-γ) and pro-inflammatory cytokine interleukin (IL) (IL-1β) and inducible nitric oxide synthase (iNOS) in chicken PBMCs were quantified in this study. Constitutive expression of IFN-β, iNOS were significantly (p < 0.01) higher in WL than that of Kadaknath and broiler birds. While, Kadaknath birds have shown significant up-regulation (p < 0.05) of IL-1β expression. This study suggests that constitutive expression of immune related genes vary with different breeds and can contribute to the differential disease resistance among these breeds.
INTRODUCTION
Immune system is critical for living organisms to combat invading pathogens, which is divided into innate and adaptive systems in vertebrates. Innate immune system, which can act immediately after the entry of the pathogens, is present across different species from insects to mammals. Adaptive immune system is mediated through high affinity rearranged receptors present on B and T lymphocytes. Innate and adaptive immunity is interlinked and need help of each others to execute the functions (Akira et al., 2001; Janeway and Medzhitov, 2002). Immune system is mediated by cellular interactions and molecules secreted by them play a major role in deciding the type and magnitude of immune response. Cytokines are central regulator of immune responses as it mediates cell signaling to produce different responses. Cells produce wide range of cytokines and it varies with respect to cells and pathogens. Based on their inherent property, cytokines represent promising candidates for treating infections that are difficult to control by antibiotics (Dinarello, 2000; Min et al., 2001; Li and He, 2004). By analyzing the constitutive expression level of immune related genes such as IFN-α, IFN-β, IFN-γ, IL-1β, TNF-α, IL-2, IL-4 and iNOS etc., prediction of immune status of the host is possible.
In this report we analyzed the basal level expression of different immune related genes viz., IFN-β and IFN-γ (antiviral), pro-inflammatory (IL-1β) and iNOS in PBMCs (ex vivo) of White Leghorn (WL), Broiler and Kadaknath breeds of chicken.
MATERIALS AND METHODS
Experimental birds
Ten day-old chicks from each WL Specific Pathogen Free (SPF) origin, Kadaknath and Broiler breeds were maintained in the Immunology section animal shed, Indian Veterinary Research Institute (IVRI), Izatnagar. Birds were provided with ad libitum feed and water and maintained following standard management practices.
Kits
Ribozol™ (AMRESCO®, USA), RevertaidTM First Strand cDNA Synthesis Kit (Thermo Scientific, USA), QuantiTect SYBR Green quantitative Real time PCR Master Mix (2X) (Qiagen, Netherlands) were procured and used for this study.
Primers
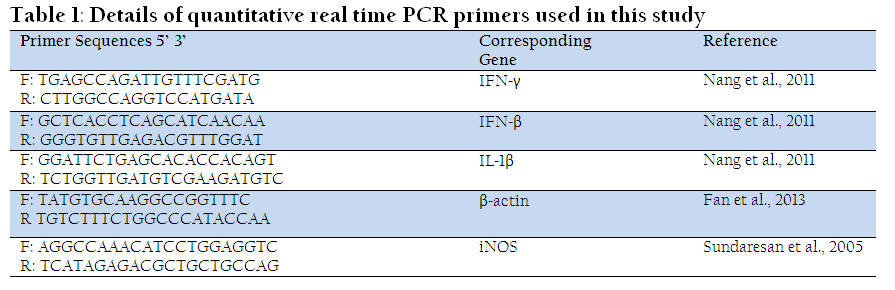
Published oligonucleotide primers were selected from earlier reports for various chicken genes viz. β-actin, IFN-β, IFN-γ, IL-1β, and iNOS. These primers were got synthesized at M/S integrated DNA technologies, USA. The details of primers used in this study are shown in Table 1.
Collection of Samples
At the age of 15 days, blood samples were collected with anticoagulant heparin (20 IU/ml) from three birds in each breed and were used for isolation of Peripheral blood mononuclear cells (PBMCs).
Isolation of Chicken PBMCs
Chicken PBMCs were isolated from blood as previously described (He et al., 2003). Blood samples with heparin were layered over Ficoll Histopaque (Sigma chemical co., USA) with the density 1.077 g/mL in equal volumes and kept for centrifugation at 400xg for 45 minutes for phase separation. Following centrifugation, the whitish cloudy interface layer containing the PBMCs was aspirated carefully through micropipette. PBMCs were washed with sterile phosphate buffered saline (PBS, pH 7.2) twice and the cell pellet was stored in 1 ml Ribozol at – 80°C until further use.
RNA extraction and cDNA Synthesis
Total RNA was extracted from PBMCs using Ribozol™ reagent as per manufacturer’s guidelines and eluted in 20 μL of nuclease free water. RNA purity and quantity were determined by Nano-drop® Spectrophotometer Analyzer at OD260 versus OD280. Using RevertaidTM First Strand cDNA Synthesis Kit, total RNA (2 μg) was reverse transcribed to cDNA with oligo-dT primers as per manufacturer’s protocol and cDNA was stored at -20°C until further use.
Real time quantitative PCR analysis
SYBR green-based quantitative Real time PCR was performed using CFX96 Touch™ (Bio-Rad). Each sample was run in duplicate. Total reaction volume was 20 μL containing QuantiTect SYBR qPCR master mix (2X) 10 μL, template cDNA 2μL (1:10 diluted with nuclease free water), gene specific forward primer 0.5 μL, reverse primer 0.5 μL and nuclease free water 7 μL. Real time PCR conditions were one cycle at 95°C for 5 min, 40 cycles at 94 °C for 45 sec, 60°C for 45 sec, 72°C for 45 sec, and 94°C for 30 sec one cycle, then 55-94°C ramp for the melt curve analysis of the PCR products for assessing the amplicon specificity. All the samples were carried out in triplicate on the same plate. The cycle at which the sample amplicon reporter dye concentration crossed a preset threshold was recorded as the cycle threshold (Ct) value. The data of quantitative real time PCR were analyzed by 2-ΔΔCt method (Pfaffl, 2001) to derive the relative fold change in mRNA expression of different genes studied. Expression levels of the above mentioned different genes were calculated in WL and Kadknath breed of birds relative to the expression of the β-actin gene and expressed as n-fold increase or decrease relative to the samples of broiler birds.
Data analysis
All the results were analyzed using the statistical software SPSS™ 20.0 (IBM, Corp. USA) and One-way analysis of variance (ANOVA) was used to determine statistical differences in mean values. All the values were considered statistically significant if p < 0.01(**) and p < 0.05(*).
RESULTS
Constitutive expression analysis of immune response genes by quantitative real time PCR method between breeds
Constitutive expression of immune related genes in PBMCs (ex vivo) were analyzed viz. IFN-β, IFN-γ, IL-β and iNOS in WL and Kadaknath breed of birds relative to broiler birds and presented in Fig.1. IFN-β expression was significantly different (p < 0.01) between all the three breeds. The highest IFN-β expression of 1.61 fold was observed in WL. IFN-γ expression showed statistically significant (p < 0.01) difference between WL and Kadaknath but not with broiler birds and the maximum fold was 1.52±0.29 in WL birds. Kadaknath birds showed significantly higher IL-1β expression than broiler and WL. IL-1β expression was reached to a maximum fold 1.67±0.21 in Kadaknath. iNOS expression showed significant difference (p < 0.05) between all the three breeds. In addition, iNOS expression was highest with a fold increase of 1.74±0.22 in WL.
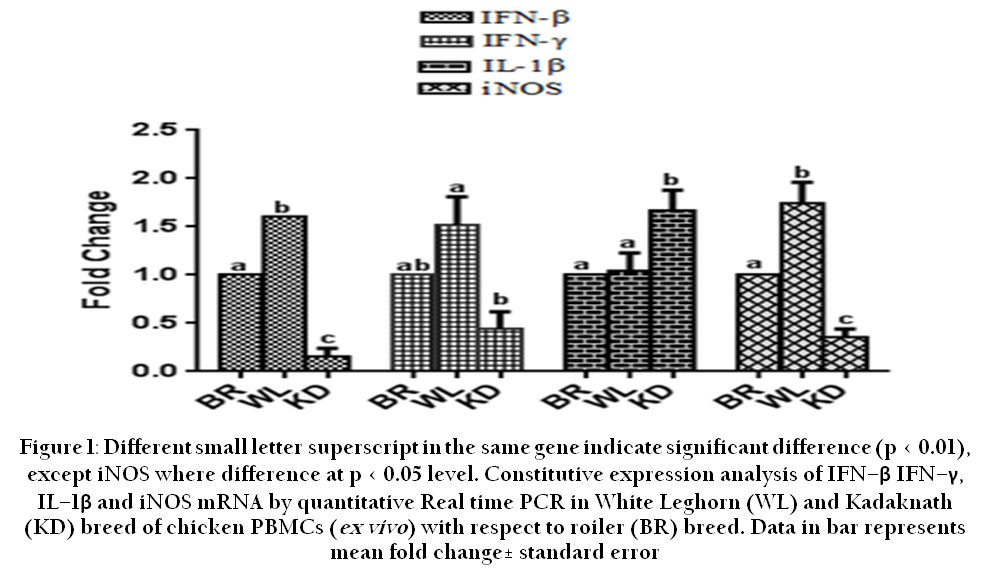
Figure 1: ifferent small letter superscript in the same gene indicate significant difference (p < 0.01), except iNOS where difference at p < 0.05 level
DISCUSSION
Most part of peripheral blood mononuclear cells comprises of monocytes/macrophages, and lymphocytes. They act as major source for different cytokines, by which they have major role in immune responses in the host. Type I interferons such as IFN-α and IFN-β both show similar activity during viral infection i.e induction of resistance to virus infection and increased expression of major histocompatibility complex (MHC) class I. Interferons after binding to target cells stimulate different types of effector genes that are responsible for antiviral state (Goodburn et al., 2000). In our study constitutive expression of IFN-β showed significant difference (p < 0.01) between WL and Kadaknath and between broilers. Of these three types of birds, WL breed showed significantly (p < 0.01) higher fold i.e. 1.61 than that of other breeds. This indicates that WL has comparatively more antiviral immune response than that of other breeds studied.
Nitric oxide is an important mediator of intra and intercellular signaling and it is diverse in bodily functions. It takes part in both antimicrobial and antitumor activity (Einestein et al., 2001). Nitric oxide is produced by a family of enzymes called as nitric oxide synthases (NOS), which is of three types based on tissue type viz. eNOS (endothelial NOS), iNOS (inducible NOS), nNOS (neuronal NOS). Of these, inducible nitric oxide synthase (iNOS), an immune defense enzyme mainly activated by PRRs (pattern recognition receptors) signaling pathways and it catalyzes the reaction of nitric oxide conversion from L-arginine (Aktan, 2004). In our study constitutive expression of iNOS was significantly higher (p < 0.01) in WL, and it was found to be significantly different (p < 0.01) between Kadaknath and WL birds. These results suggest that WL has constitutively superior nitric oxide production level than that of other breeds. Earlier study involves comparison between Aseel and Kadaknath, in which Kadaknath showed significant higher level of serum nitric oxide level than that of Aseel breed (Kannaki et al., 2011). However, this study did not compare between Kadaknath and Aseel for constitutive expression of iNOS. Studies revealed that expression of iNOS is under the genetic influence (Dil and Qureshi, 2002), which was observed in our study.
IFN-γ is a type II IFN, chiefly produced by activated T lymphocytes (cytotoxic & helper), NK cells, B cells, and professional antigen presenting cells. It has immunomodulatory effects because it induces Th1 biased immune response, which is evidenced by stimulation of Th1 differentiation by this gene expression (Gessani and Belardelli, 1998; Yoshimoto et al., 1998; Frucht et al., 2001; Schroder et al., 2004). Constitutive expression of IFN-γ showed significant difference (p < 0.01) between WL and Kadaknath.
IL-1β up-regulation leads to induction of pro-inflammatory cytokines and adhesion molecules. IL-1β secreted mainly by macrophage/monocytes,and dendritic cells (Klasing and Peng, 1987; Beuscher et al., 1990; Weining et al., 1998) . Constitutive expression of IL-1β was significant (p < 0.05) in Kadaknath in comparison with broiler. In addition, it was significantly (p < 0.05) different between WL and Kadaknath. However, there was no significant difference occurred between broiler and WL. Expression of IL-1β in Kadaknath was significantly higher than other breeds, which indicates comparatively enhanced inflammatory activity in indigenous breeds. . Similar to our study, PBMCs from Kadaknath and Aseel breeds of chicken were analyzed for different TLRs constitutive expression and showed that Kadaknath has significantly higher expression in TLR3, 15 and 21 while Aseel breed has showed higher constitutive expression only in TLR4. Further they suggested that Kadaknath has better immunecompetence than that of Aseel breed (Patel et al., 2013).
In conclusion, the present study reports the variation in expression of immune related genes in ex vivo PBMCs of different breeds viz. Broiler, WL and Kadaknath. The variation in expression of these genes might contribute to the differences in disease resistance among these commercial and indigenous chicken breeds. Future studies could be aimed to study constitutive cytokine expression level in different organs.
REFERENCES
Akira S, Takeda K, Kaisho T (2001). Toll-like receptors: critical proteins linking innate and acquired immunity. Nat. Immunol. 2: 675 - 680.
http://dx.doi.org/10.1038/90609
PMid:11477402
Aktan F (2004). iNOS-mediated nitric oxide production and its regulation. Life Sci. 75: 639 - 653.
http://dx.doi.org/10.1016/j.lfs.2003.10.042
PMid:15172174
Beuscher HU, Gunther C, Rollinghoff M (1990). IL-1 beta is secreted by activated murine macrophages as biologically inactive precursor. J. Immunol. 144: 2179 - 2183.
PMid:2313092
Dil N, Qureshi MA (2002). Involvement of LPS related receptors and nuclear factor Kappa-B indifferential expression of inducible nitric oxide synthase in chicken macrophages from different genetic backgrounds. Vet. Immunol. Immunopathol. 88: 149 - 161.
http://dx.doi.org/10.1016/S0165-2427(02)00153-8
Dinarello CA (2000). Proinflammatory cytokines. Chest. 118: 503 - 508.
http://dx.doi.org/10.1378/chest.118.2.503
PMid:10936147
Eisenstein TK (2001). Implications of Salmonella induced nitric oxide (NO) for host defense and vaccines: NO, an antimicrobial, antitumor, immunosuppressive and immunoregulatory molecule. Microbes. Infect. 3: 1223 - 1231.
http://dx.doi.org/10.1016/S1286-4579(01)01482-4
Fan YF, Hou ZC, Yi GQ, Xu GY,Yang N (2013). The sodium channel gene family is specifically expressed in hen uterus and associated with eggshell quality traits. BMC Gen. 14: 90.
http://dx.doi.org/10.1186/1471-2156-14-90
PMid:24059973 PMCid:PMC3851161
Frucht DM, Fukao T, Bogdan C, Schindler H, O'Shea JJ, Koyasu S (2001). IFN-γ production by antigen-presenting cells: mechanisms emerge. Trends Immuno. 22: 556 - 560.
http://dx.doi.org/10.1016/S1471-4906(01)02005-1
Gessani S, Belardelli F (1998). IFN-γ Expression in Macrophages and Its Possible Biological Significance. Cytokine Growth F. R. 9: 117 - 123.
Goodbourn S, Didcock L, Randall RE (2000). Interferons: cell signalling, immune modulation, antiviral response and virus countermeasures. J. Gen.Virol. 81: 2341 - 2364.
PMid:10993923
He H, Crippen TL, Farnell MB, Kogut MH (2003). Identification of CpG oligodeoxynucleotide motifs that stimulate nitric oxide and cytokine production in avian macrophage and peripheral blood mononuclear cells. Develop. Comp. Immunol. 27: 621 – 627.
http://dx.doi.org/10.1016/S0145-305X(03)00013-2
Janeway Jr CA, Medzhitov R (2002). Innate immune recognition. Annu. Rev. Immunol. 20: 197 - 216.
http://dx.doi.org/10.1146/annurev.immunol.20.083001.084359
PMid:11861602
Kannaki TR, Reddy MR, Murugesan S (2011). Toll-like receptor mRNA expression, iNOS gene polymorphism and serum nitric oxide levels in indigenous chickens. Vet. Res. Comm. 35: 321 - 327.
http://dx.doi.org/10.1007/s11259-011-9472-z
PMid:21607608
Klasing KC, Peng RK (1987). Influence of cell sources, stimulating agents, and incubation conditions on release of interleukin-1 from chicken macrophages. Dev. Comp. Immunol. 11: 385 - 394.
http://dx.doi.org/10.1016/0145-305X(87)90082-6
Li MC, He SH (2004). IL-10 and its related cytokines for treatment of inflammatory bowel disease. World J. Gastroentero. 10: 620 - 625.
PMid:14991925
Min W, Lillehoj HS, Burnside J, Weining KC, Staeheli P, Zhu JJ (2001). Adjuvant effects of IL-1β, IL-2, IL-8, IL-15, IFN-α, IFN-γ, TGF-β4 and lymphotactin on DNA vaccination against Eimeria acervulina. Vaccine. 20: 267 - 274.
http://dx.doi.org/10.1016/S0264-410X(01)00270-5
Nang NT, Lee JS, Song BM, Kang YM, Kim HS, Seo SH (2011). Induction of inflammatory cytokines and toll-like receptors in chickens infected with avian H9N2 influenza virus. Vet. Res. 42: 10 - 1186.
http://dx.doi.org/10.1186/1297-9716-42-64
PMid:21592354 PMCid:PMC3114738
Patel MV, Kumar P, Das H, Dadawala A, Chauhan HC, Chandel BS, Kher HN (2013). Evaluation of the constitutive expression levels of ch-TLR 3, ch-TLR 4, ch-TLR 15 and ch-TLR 21 genes in the Peripheral Blood Mononuclear Cells of native Indian poultry breeds, Aseel and Kadaknath.Vet.World. 6: 568 - 572.
http://dx.doi.org/10.5455/vetworld.2013.568-572
Schroder K, Hertzog PJ, Ravasi T, Hume DA (2004). Interferon-γ: an overview of signals, mechanisms and functions. J. Leukocyte Biol. 75: 163 - 189.
http://dx.doi.org/10.1189/jlb.0603252
PMid:14525967
Sundaresan NR, Ahmed KA, Saxena VK, Sastry KVH, Saxena M, PramodAB, Nath M, Singh KB, Rasool TJ, Devroy AK, Singh RV (2005). Differential expression of inducible nitric oxide synthase and cytokine mRNA in chicken lines divergent for cutaneous hypersensitivity response. Vet. Immunol. Immunopathol. 108: 373 - 385.
http://dx.doi.org/10.1016/j.vetimm.2005.06.011
PMid:16099515
Weining KC, Sick C, Kaspers B, Staeheli P (1998). A chicken homolog of mammalian interleukin‐1β: cDNA cloning and purification of active recombinant protein. Eur. J. Biochem. 258: 994 - 1000.
http://dx.doi.org/10.1046/j.1432-1327.1998.2580994.x
PMid:9990317
Yoshimoto T, Takeda K, Tanaka T, Ohkusu K, Kashiwamura SI, Okamura H, Nakanishi K (1998). IL-12 up-regulates IL-18 receptor expression on T cells, Th1 cells, and B cells: synergism with IL-18 for IFN-γ production. J.Immunol. 161: 3400 - 3407
PMid:9759857

