
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences. 1 (4S): 30 – 36Special Issue–4 (Progress in Research on Viruses and Viral Diseases)
Vp5 Gene Based Molecular Characterization of Bluetongue Virus 9 from South India
Koushlesh Ranjan, Gaya Prasad,, Pawan Kumar, Prasad Minakshi*
-
Department of Animal Biotechnology, LLR University of Veterinary and Animal Sciences, Hisar, Haryana, India, 125004
*Corresponding author:[email protected]
ARTICLE CITATION:
Ranjan K, Prasad G, Kumar P and Minakshi P (2013). Vp5 gene based molecular characterization of bluetongue virus 9 from South India. Adv. Anim. Vet. Sci. 1 (4S): 30 – 36.
Received: 2013–09–10, Revised: 2013–11–15, Accepted: 2013–11–16
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/137/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Bluetongue virus (BTV) is a prototype species of the genus Orbivirus within the family Reoviridae that causes Bluetongue disease (BT) in domestic as well as wild ruminants. The BTV isolate Ong5/06/Ind was isolated from sheep in Andhra Pradesh state in 2006. The isolate was confirmed as BTV based on characteristics CPE in BHK21 cell culture, 3:3:3:1 migration pattern of viral nucleic acid in RNA–PAGE and 366bp amplicon with group specific ns1 gene based PCR. VP5 gene based serotype specific primer produced specific PCR amplicon of 823bp corresponding to BTV serotype 9. Nucleotide sequence analysis revealed 97% identity with other Indian and European BTV–9 isolates, more than 90% with Asian and, only 68.8% with South African isolates. However, based on deduced amino acids, the identity was more than 97% with Indian, Asian and European and, only 73.4% with South African BTV–9 isolates. The in–silico restriction enzyme analysis (REA) with AflIII, BtrI and HindII showed a common pattern of restriction sites in Indian and European isolates at 1152, 1153 and 591, respectively. The lack of any of these restriction sites in African and other Asian isolates revealed the European origin of the study isolate. Similarly, the lack of BsmAI restriction site in Ong5/06/Ind isolate (Accession no. JF969307), differentiate it from European, Asian and South Africa isolates. The sequence analysis revealed that Ong5/06/Ind, Asian and European BTV–9 isolates were placed in nucleotype ‘C’ while South African isolates were placed in nucleotype ‘B’.
INTRODUCTION
Bluetongue (BT) is an insect–borne viral disease of domestic and wild ruminants caused by bluetongue virus (BTV), belonging to genus Orbivirus and family Reoviridae. BTV is a non enveloped dsRNA virus. The genome of BTV consists of 10 segments dsRNA encoding 7 structural (VP1 to VP7) and 4 non structural proteins (NS1, NS2, NS3/NS3a and NS4) (Ratinier et al., 2011; Belhouchet et al., 2011). The segment 2 (VP2 gene) is major serotype specific along with minor serotype specific segment 6 (VP5 gene). Genetic analysis of various BTV serotypes and strains has revealed the existence of distinct Eastern and Western topotypes associated with specific geographical origin of virus (Maan et al., 2010). In addition, VP2 and VP5 nucleotypes have also been proposed based on nucleotide sequence relatedness, and this roughly correlates with closely related serotypes as well as cross–neutralization patterns (Maan et al., 2010). BTV is transmitted by biting midges (Culicoides spp.) and can infect all species of ruminant (Maclachlan and Guthrie, 2010) especially small ruminants such as sheep. Clinical signs of BTV infection are more severe and confined to sheep and white–tailed deer (Darpel et al., 2007).
Due to rapid evolutionary changes in genome through reassortment and mutations, BTVs are consistently evolving new serotypes globally. Twenty–four distinct serotypes of the BTV have been reported around the globe (Mertens et al., 2004). However two more serotypes BTV25 from Switzerland (Hofmann et al., 2008) and BTV26 from Kuwait (Maan et al., 2011) have been isolated recently.
Since India is endemic for BTV, a total of 21 different serotypes have been reported based on virus isolation and serum neutralization test (Prasad et al., 2009). Recently, BTV serotype 21 has been reported from India (Jordar et al., 2009). The complete genome sequence of BTV16 of goat origin from India has been done recently (Minakshi et al., 2012). BTV is spreading in newer areas continuously; seroprevalence of BTV has been reported from Kerala state (Ravishankar et al., 2005). However, most of the studies were done on VP2 gene basis but in present study VP5 gene based serotype confirmation and molecular characterization of one of BTV isolate is done.
MATERIALS AND METHODS
Virus Sample Preparation
The virus was isolated during an outbreak in a sheep flock (of Nellore breed) in Andhra Pradesh state in 2006. The sample was designated as a new isolate Ong5/06/Ind since it was not reported earlier. The virus was propagated in BHK–21 cell line. After appearance of about 75% cytopathetic effect (CPE) at 6th Passage in infected BHK–21 cell line, the virus was harvested. The dsRNA viral nucleic acid was extracted from cell culture grown virus by Trizol® (Sigma, USA) method as per manufacturer’s instructions.
RNA Poly Acrylamide Gel Electrophoresis (RNA–PAGE) of Viral Nucleic Acid
The viral nucleic acid was subjected for 8% RNA–PAGE to determine the specific migration pattern of BTV specific nucleic acid. To visualize the nucleic acid segments RNA–PAGE was allowed for silver staining (Svensson et al., 1986) (data not shown).
Oligonucleotide primers
To confirm the new isolate as BTV, a serogroup specific NS1 gene based primer of amplicon size 366 bp was used (Kovi et al., 2005). The primer pair P1 (443–462nt) – 5’ CGGGAGGAGGCGGAACAGGA3’ and P2 (1246 –1265nt) – 5’CGTGCCAGTGGGCAGCAAGA3’ specific to VP5 gene of BTV–9 serotype was designed using sequences available in GenBank with the help of Primer Blast software (http://www.ncbi.nlm.nih.gov/tools/primer–blast/). The specific amplicon size of this primer pair was 823bp. Specificity of the primer was tested by BLASTN similarity search in GenBank database.
Polymerase Chain Reaction (PCR)
The cDNA was prepared from viral nucleic acid using random primers (decamer) (Ambion, USA) and moloney murine leukemia virus–reverse transcriptase (Mo–MuLV–RT) enzyme (Promega) as per manufacturer’s instruction. The cDNA was used for NS1 gene based PCR to confirm the isolate as BTV using standard protocol (Kovi et al., 2005). The cDNA was subsequently used for amplification of VP5 gene using primers specific to all the BTV serotypes including primers P1 and P2. The amplification was carried out in 20 µL reaction mixture containing 2 µL cDNA, 3% DMSO, 20 µM of primers along with 0.4 µL of 10mM dNTPs mix, 4 µL 5X HF buffer and 0.4 U (2 U/µL) phusion high– fidelity DNA polymerase (Finnzymes, Finland) in thermalcycler (Biorad iCycler, USA). The thermal condition for primer P1 and P2 was set as, initial denaturation for 3 min at 98oC, followed by 35 cycles for 10 sec denaturation at 98oC, 20 sec primer annealing at 57oC and 30sec primer extension at 72oC. Final primer extension was carried out at 72oC for 10 min. The PCR products were analysed by agarose gel electrophoresis using 1% agarose (Sigma, USA).
Nucleotide Sequencing and Data Analysis
The VP5 gene specific PCR amplicon was sequenced using Genetic Analyser ABI PRISM TM 3130 XL. The sequence data obtained was analysed using online software BLASTN+ 2.2.28 (http://blast.ncbi.nlm.nih.gov/) (Zhang et al., 2000). The nucleotide sequence was translated to deduced amino acid sequence according to its open reading frame. BioEdit v7.2.3 software was used for calculation of percent nucleotide and amino acid sequence identity of VP5 gene of Ong5/06/Ind isolate (Accession no. JF969307) with BTV9 global isolates and all the 26 BTV serotypes. MEGA5 (Tamura et al., 2011) software was used for nucleotide as well as amino acid based phylogenetic analysis. The in silico REA of VP5 gene of virus isolate used in present study along with other global isolates were carried out using Restriction Mapper version 3.0 software available at http://www.restrictionmapper.org/.
RESULTS
The Ong5/06/Ind isolate (Accession no. JF969307) produced BTV specific CPE such as detachment, aggregation of the cells, foamy degeneration of the cells etc. in BHK–21 cell culture. The RNA–PAGE analysis showed BTV specific migration pattern (3:3:3:1) of viral dsRNA (data not shown). The Ong5/06/Ind isolate produced an expected amplicon size of 366bp with group specific NS1 gene based PCR (Figure 1). The VP5 gene of Ong5/06/Ind isolate was successfully amplified using primers P1 and P2 (specific to BTV9) as evidenced by expected PCR amplicon size of 823bp in agarose gel electrophoresis (Figure 2).
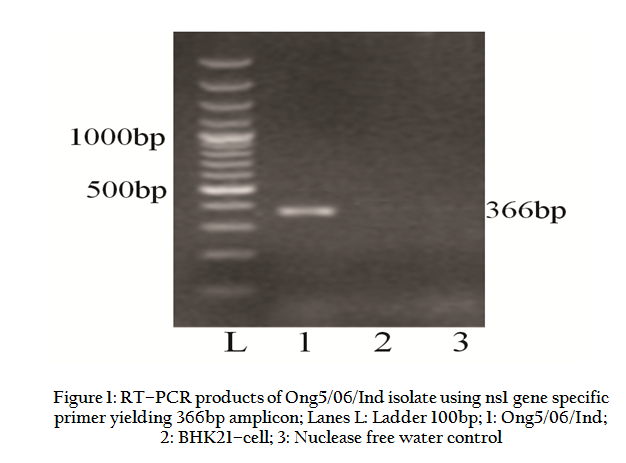
Figure 1: RT–PCR products of Ong5/06/Ind isolate using ns1 gene specific primer yielding 366bp amplicon
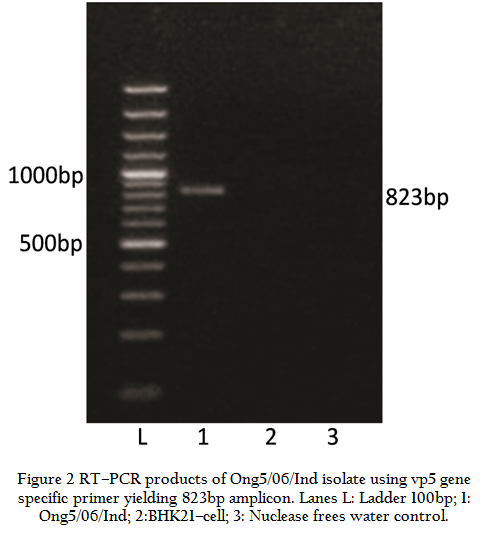
Figure 2: RT–PCR products of Ong5/06/Ind isolate using vp5 gene specific primer yielding 823bp amplicon
Similarly, Ong5/06/Ind isolate was also subjected to VP5 gene specific primers of remaining BTV serotypes. However, it did not give any amplification with VP5 gene specific primers of remaining serotypes (data known shown). The VP5 gene PCR amplicon was sequenced and sequence data was deposited to GenBank with accession no. JF969307. Ong5/06/Ind isolate showed maximum nucleotide (>97 %) as well as deduced amino acid (>98%) sequence identity with European and other Indian isolates (Table 1). The phylogenetic analysis of Ong5/06/Ind isolate (Accession no. JF969307) along with representative BTV sequences from different parts of the world were done using Mega5 software (Tamura et al., 2011). The nucleotide as well as deduced amino acid sequence based phylogenetic analysis of partial VP5 gene revealed that Ong5/06/Ind isolate (Accession no. JF969307) form a close cluster with European and other Indian isolates of BTV9 (Figure 3 and 4). The phylogenetic analysis of VP5 gene of all the 26 BTV serotypes revealed that Ong5/06/Ind isolate along with other Indian and European BTV9 isolates was placed in nucleotype ‘B’ whereas South African BTV9 isolates were placed in nucleotype ‘C’ (Figure 5).
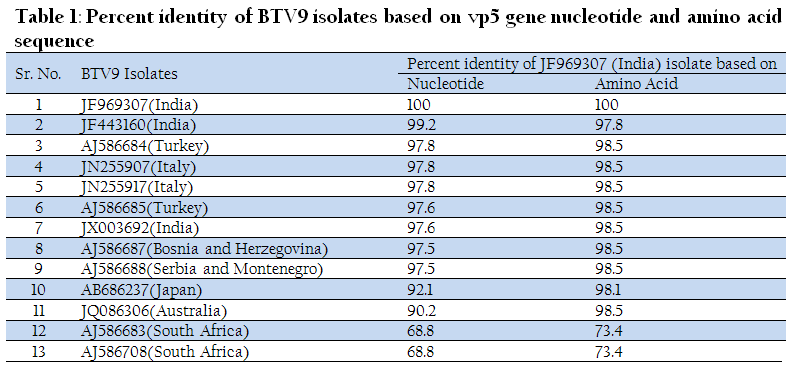
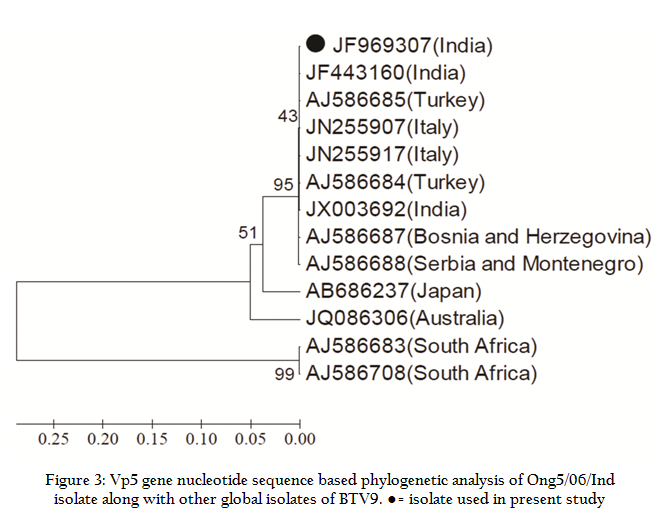
Figure 3: Vp5 gene nucleotide sequence based phylogenetic analysis of Ong5/06/Ind isolate along with other global isolates of BTV9
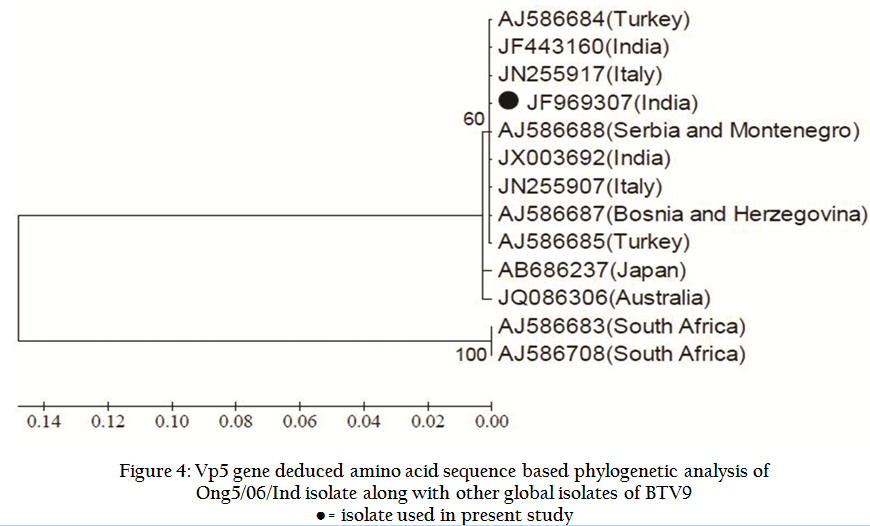
Figure 4: Vp5 gene deduced amino acid sequence based phylogenetic analysis of Ong5/06/Ind isolate along with other global isolates of BTV9
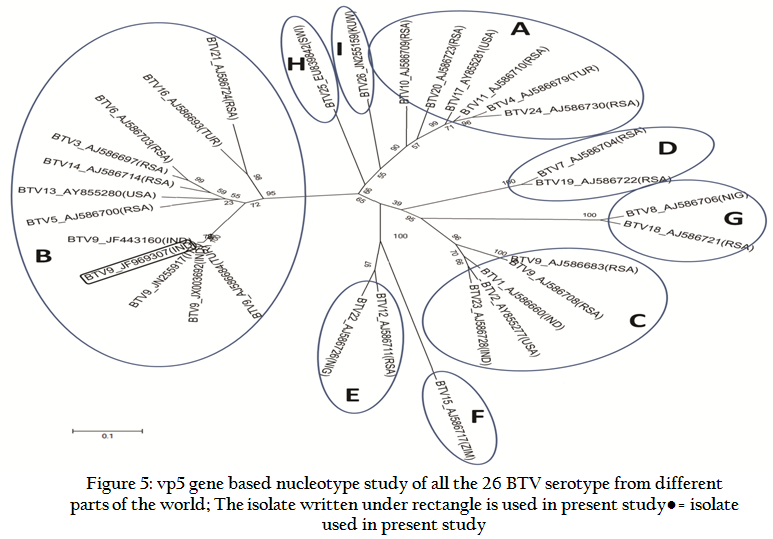
Figure 5: vp5 gene based nucleotype study of all the 26 BTV serotype from different parts of the world; The isolate written under rectangle is used in present study
The in silico REA of VP5 gene with enzymes BtrI, HindII and AflIII revealed that Ong5/06/Ind isolate share a common pattern of restriction sites with that of European and other Indian isolates (Table 2). However, restriction enzyme BsmAI showed genetic diversity of Indian isolates with other global isolates of BTV9. The VP5 gene based nucleotide and deduced amino acid sequence identity of all the 26 BTV serotypes was calculated using Bioedit v7.2.3 software programme. It was observed that Ong5/06/Ind isolate along with other Indian and European isolates had shown maximum identity with nucleotype ‘B’ serotypes and south African BTV9 isolate had shown maximum identity with Nucleotype ‘C’ serotypes (Table 3).
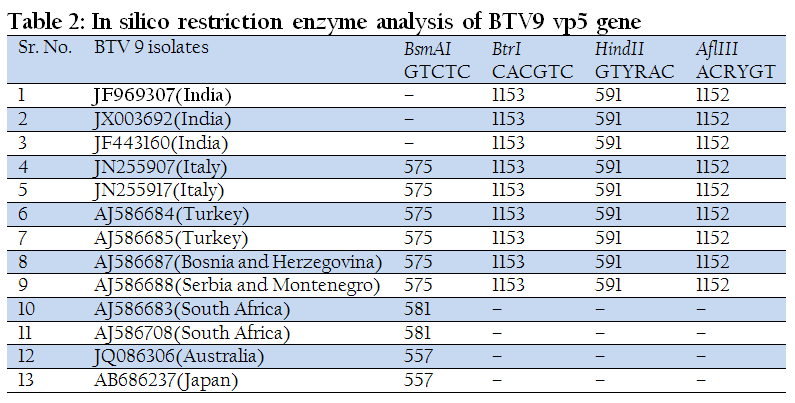
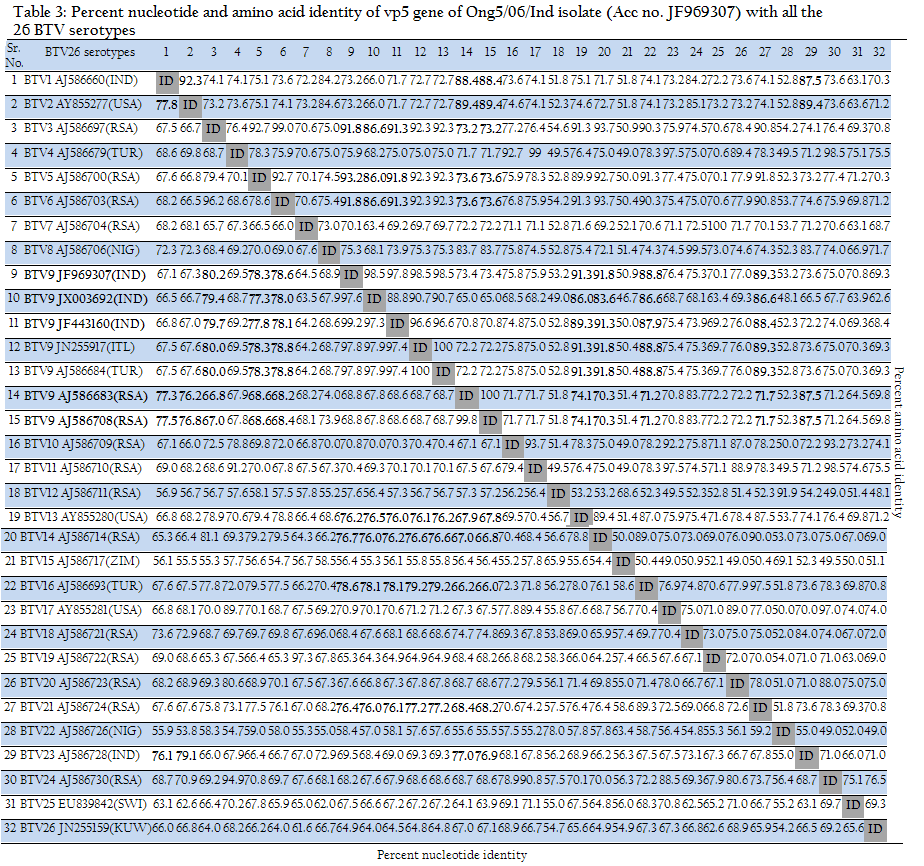
Table 3: Percent nucleotide and amino acid identity of vp5 gene of Ong5/06/Ind isolate (Acc no. JF969307) with all the 26 BTV serotypes
DISCUSSION
BT is an endemic disease in India for domestic and wild ruminants. A large number of BTV serotypes have been reported from India and BTV9 is one of the common serotype prevalent in country. BT is reported from various states of India such as Himachal Pradesh, Uttar Pradesh, Gujarat, Maharashtra and Andhra Pradesh (Prasad et al., 2009). VP5 gene based molecular characterization and phylogenetic relationship of BTV1 of Indian isolates (Manjunath et al., 2010) and reference strains of all 24 serotypes (Singh et al., 2004) have been reported earlier. The prevalence of BTV–9 was reported earlier from southern part of India (Rao et al., 2012). The present investigation was carried out for VP5 gene based serotype confirmation of an Indian BTV isolate and its comparison with other BTV isolates from India and global origin.
The BTV specific characteristics CPE in BHK–21 cell culture, migration pattern in 8% RNA–PAGE and an expected amplicon size with group specific ns1 gene based RT–PCR confirmed the samples as BTV. The VP5 gene nucleotide based phylogenetic analysis using neighbour joining (NJ) method of MEGA5 software revealed that Ong5/06/Ind isolate formed a closer cluster with European and other Indian isolates. However it formed a minor cluster with Japan and Australia isolates. South Africa isolates were related distantly to it. The similar result was observed with deduced amino acid based phylogenetic analysis.
The REA of full or partial length gene is useful in differentiating between serotypes and within isolates of the same serotype. The in–silico REA was done with BtrI, AflIII, HindII and BsmA1 restriction enzymes. The in–silico REA revealed the single restriction site for restriction enzymes BtrI, AflIII and HindII at nucleotide 1153, 1152 and 591 respectively in all the Indian and European isolates including isolates used in present study. However, these enzymes did not show any restriction sites in Japan, Australia and South Africa isolates. Similarly, REA with BsmA1 revealed single restriction site at nucleotide 575 in all the European isolates. The same BsmA1 restriction site was present in South Africa at 581 and in Australia and Japan isolates at nucleotide 557. The restriction enzymes BtrI, AflIII and HindII showed a common restriction pattern in Indian and European BTV9 isolates. It indicated the European origin of our Indian isolates. However, BsmA1 can be used for differentiation of Indian BTV9 isolates with European, South Africa, Japan and Australia isolates.
The phylogenetic analysis and in–silico REA indicated that Ong5/06/Ind isolate was of European origin. The analyses of partial vp5 gene sequence revealed that there is no significant divergence among Indian BTV9 isolates. They show only 0–0.4% nucleotide and 0–0.7% deduced amino acid based diversity. It indicates the common origin of these viruses. The Segment–6 sequences of the all the 26 different BTV serotypes cluster as 8 distinct evolutionary lineages, called as segment 6 nucleotypes (nucleotypes A–I) (Maan et al., 2011). The nucleotype ‘B’ includes serotype 3, 5, 6, 13, 14, 16 and 21whereas nucleotype ‘C’ includes serotype 1, 2, 9 and 23 (Maan et al., 2010). The grouping of the different BTV serotypes within these 9 nucleotypes also reflects the serological relationships between them i.e. low–level cross–reactions between different BTV serotypes. The identity percentage for nucleotides and amino acids within a nucleotype was reported as 76–100% and 86–100% for nucleotides and amino acids respectively. However, the identity percentage for nucleotides and amino acids between different nucleotypes and serotypes was reported as 56.7–76% and 54.2–85.9% respectively (Maan et al., 2010).
The Ong5/06/Ind isolate along with other Indian and European BTV9 isolates showed maximum 76–80.2% nucleotide and 88–93.2% amino acids, identity with nucleotype ‘B’ serotypes (serotype 3, 5, 6, 13, 14, 16 and 21). This placed our Ong5/06/Ind isolate along with other Indian and European BTV9 isolates in nucleotype ‘B’ (Fig. 5). However, the BTV9 isolates from South Africa had shown maximum 76.2–77.5% nucleotide and 87.5–89.4% amino acid identity with nucleotype ‘C’ serotypes (serotypes 1, 2 and 23). This placed South African BTV9 in nucleotype ‘C’ as reported earlier (Maan et al., 2010). However, all the Indian and European BTV9 isolates had shown 66.5–69.3% nucleotide and 66.0–73.6% amino acid, identity with nucleotype ‘C’ serotypes. Similarly, South African BTV9 isolates had shown 66–68.6% nucleotide and 70.3–74.1% amino acid identity with the nucleotype ‘B’ serotypes (Table 3). The above discussion clearly indicated the existence of at least two segment–6 nucleotype in BTV9 globally.
CONCLUSION
A new BT virus isolated from Andhra Pradesh state had been confirmed as BTV serotype 9 based on vp5 gene specific PCR and nucleotide sequencing technique. Our vp5 gene based molecular analysis confirmed the virus isolate is of European origin. The segment–6 analysis of all the 26 serotypes revealed presence of at least two nucleotype in BTV9. The vp5 gene based phylogenetic analysis and REA tools could be effectively used for establishing phylogenetic relationship and differentiation of Indian isolates from rest of the global BTV9 isolates. The vp5 gene specific primer tested in this study can be used for identifying the BTV9 serotype prevalent in endemic region, and suitable vaccine candidate can be identified for controlling the disease.
ACKNOWLEDGEMENT
The research was funded by Indian council of Agricultural Research, New Delhi under all India network programme on Bluetongue. The authors are thankful to Department of Animal Biotechnology, LLR University of Veterinary and Animal Sciences, Hisar for providing infrastructural facility.
CONFLICT OF INTEREST
We declare no conflict of interest.
REFERENCES
Belhouchet M, Mohd Jaafar F, Firth AE, Grimes JM, Mertens PPC and Attoui H (2011). Detection of a fourth Orbivirus non–structural protein. PLoS ONE. 6: e25697.
http://dx.doi.org/10.1371/journal.pone.0025697
PMid:22022432 PMCid:PMC3192121
Darpel KE, Batten CA, Veronesi E, Shaw AE, Anthony S, Bachanek–Bankowska K, Kgosana L, Bin–Tarif A, Carpenter S, Muueller–Doblies UU, Takamatsu HH, Mellor RS, Mertens PPC and Oura CAL (2007). Clinical signs and pathology shown by British sheep and cattle infected with Bluetongue virus serotype 8 derived from the 2006 outbreak in northern Europe. Vet. Rec. 161:253 – 261.
http://dx.doi.org/10.1136/vr.161.8.253
PMid:17720961
Hofmann M, Renzullo S, Mader M, Chaignat V, Worwa G and Thuer B (2008). Genetic characterization of Toggenburg Orbivirus a new bluetongue virus from goats Switzerland. Emerg. Infect. Dis. 121:855 – 1861.
Jordar SN, Lodh C, Chakrabarti A, Baksi S and Aich R (2009). Isolation of Bluetongue virus serotype 15 and 21 in West Bengal, India. Vet. Rec. 165:751 – 752.
Kovi RC, Dahiya S, Minakshi and Prasad G. (2005). Evaluation of new primers based on serogroup specific genes for detection of bluetongue viruses by RT–PCR. Indian J. Microbiol. 45: 307 – 309.
Maan S, Maan NS, Nomikou K, Veronesi E, Bachanek–Bankowska K, Manjunatha BN, Houssam A and Mertens PPC (2011). Complete Genome Characterisation of a Novel 26th Bluetongue Virus Serotype from Kuwait. PLoS ONE 6: e26147. doi:10.1371/journal.pone.0026147.
http://dx.doi.org/10.1371/journal.pone.0026147
Maan S, Maan NS, van Rijn PA, van Gennip RGP, Sanders A, Wright IM, Batten C, Hoffmann B, Eschbaumer M, Oura CAL, Potgieter AC, Nomikou K and Mertens PPC (2010). Full genome characterisation of bluetongue virus serotype 6 from the Netherlands 2008 and comparison to other field and vaccine strains. PLoS ONE. 5(4): 1 – 17.
http://dx.doi.org/10.1371/journal.pone.0010323
PMid:20428242 PMCid:PMC2859060
Maclachlan NJ and Guthrie AJ (2010). Re–emergence of bluetongue, African horse sickness,and other Orbivirus diseases. Vet. Res. 41: 35.
http://dx.doi.org/10.1051/vetres/2010007
PMid:20167199 PMCid:PMC2826768
Manjunath BN, Prasad M, Maan S and Prasad G (2010). Differentiation of Indian isolates of bluetongue virus serotype 1from Australian and African isolates based on analysis of vp5 gene. Indian J. Biotech. 9:117 – 125.
Mertens PPC, Maan S, Samuel A and Attoui H (2004). Orbivirus, Reoviridae. In: Virus Taxonomy, VIIth Report of the ICTV, edited by C.M. Fauquet, M.A. Mayo, J. Maniloff, U. Desselberger, L.A. Ball (London: Elsevier/Academic Press), 466 – 483.
Minakshi P, Singh R, Ranjan K, Kumar P, Joshi CG, YKM Reddy and Prasad G (2012). Complete genome sequence of bluetongue virus serotype 16 of goat origin from India. J. Virol. 86:8337 – 8338.
http://dx.doi.org/10.1128/JVI.01128-12
PMid:22787269 PMCid:PMC3421637
Prasad G, Sreenivasulu D, Singh KP, Mertens PPC and Maan S (2009). Bluetongue in the Indian subcontinent. In: Bluetongue. (Eds. Mellor P, Baylis M and Merten P C). Elsevier Ltd., London. 167 – 195.
http://dx.doi.org/10.1016/B978-012369368-6.50012-5
PMid:19339108
Rao PP, Reddy YV, Meena K, Karunasree N, Susmitha B, Uma M, Prasad PUVS, Chaitanya P, Reddy YN and Hegde NR (2012). Genetic characterization of bluetongue virus serotype 9 isolates from India. Virus Genes. DOI 10.1007/s11262–011–0707–4.
http://dx.doi.org/10.1007/s11262-011-0707-4
Ratinier M, Caporale M, Golder M, Franzoni G, Allan K, Armezzani A, Bayoumy A, Rixon F, Shaw A and Palmarini M (2011). Identification and Characterization of a Novel Non–Structural Protein of Bluetongue Virus. PLoS Pathog. 7(12): e1002477. doi:10.1371/journal.ppat.1002477.
http://dx.doi.org/10.1371/journal.ppat.1002477
Ravishankar C, Nair GK, Mini M and Jayaprakasan V (2005). Seroprevalence of bluetongue virus antibodies in sheep and goats in Kerala State, India. Rev. Sci. Tech. Off. Int. Epiz. 24: 953 – 958.
Svensson L, Uhnoo I, Grandien M and Wadeli G (1986). Molecular epidemiology of rotavirus infections in Upsala. Sweden. 1981; disappearance of a predominant electropherotype. J. Med. Virol. 18: 101 – 111.
http://dx.doi.org/10.1002/jmv.1890180202
PMid:3005484
Singh KP, Maan S, Samuel AR, Rao S, Meyer AJ and Mertens PPC (2004). Phylogenetic analysis of bluetongue virus genome segment 6 (encoding VP5) from different serotypes. Vet. Ital. 40:479 – 483.
PMid:20422573
Tamura K, Peterson D, Peterson N, Stecher G, Nei M and Kumar (2011). MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol Biol Evol. 28:2731 – 2739.
http://dx.doi.org/10.1093/molbev/msr121
PMid:21546353 PMCid:PMC3203626
Zhang Z, Schwartz S, Wagner L and Miller W (2000). A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 7:203 – 214.
http://dx.doi.org/10.1089/10665270050081478
PMid:10890397

