
Advances in Animal and Veterinary Sciences


Review Article
Advances in Animal and Veterinary Sciences. 1 (2S): 1 – 6Special Issue-2 (Clinical Veterinary Practice –T rends)
RNA Interference in Parasites; Prospects and Pitfalls
Naduvanahalli Rajanna Sudhakar1, Haranahalli Vasanthachar Manjunathachar1*, Kumaragurubaran Karthik2, Shivani Sahu3, Marappan Gopi4, Sanganagouda Biradar Shantaveer5, Doddhadasarahalla Nanjappa Madhu6, Prem Sagar Maurya1, Keresara Hariyappa Nagaraja7, Santhosh Shinde8, Paramasivam Tamilmahan6
- Division of Parasitology, Indian Veterinary Research Institute, Izatnagar, India
- Division of Bacteriology and Mycology, Indian Veterinary Research Institute, Izatnagar, India
- Veterinary Officer, Dehra dun, Veterinary Research Institute, Izatnagar, India
- Division of Animal Nutrition Indian Veterinary Research Institute, Izatnagar, India
- Veterinary Officer, Karanataka
- Division of Veterinary Surgery and Radiology, Indian Veterinary Research Institute, Izatnagar, India
- Division of Veterinary Public Health, Indian Veterinary Research Institute, Izatnagar, India
- Division of Veterinary Obstetrics and Gynecology Indian Veterinary Research Institute, Izatnagar, India
*Corresponding author:[email protected]
ARTICLE CITATION:
Sudhakar N R, Manjunathachar H V, Karthik K , Sahu S, Gopi M, Shanthaveer S B, Nagaraja K H, Shinde S and Tamilmahan P, Madhu D N, and Maurya PS (2013). RNA interference in parasites: prospects and pitfalls. Adv. Anim. Vet. Sci. 1 (2S): 1 – 6.
Received: 2013–09–16, Revised: 2013–10–10, Accepted: 2013–10–11
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/113/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Parasites comprise of helminthes, protozoa and arthropods which are the most complex and well adopted organisms in the host causing chronic illness in the animals resulting in considerable economic losses in the form of decreased production and loss of condition. Until today the effective control of these organisms is not possible due to the emergence of antiparasitic drugs resistance and availability of very few successful vaccines against them. By the advent of the RNA interference (RNAi) technique in late 19th century it was hoped that dream of effective control of parasites were made possible. However, after 10–15 years of RNAi research the fruitful results are still awaited. The RNAi is a process of introduction of double–stranded RNA (dsRNA) into some cells or organisms, resulting in degradation of its homologous mRNA. The dsRNAs are processed into short interfering RNAs (siRNAs) that subsequently bind to the RNA–induced silencing complex (RISC), causing degradation of target mRNAs. The sequence–specific ability of RNAi to silence target genes has been extensively used to study gene functions and to control disease pathogens and vectors. Thus, RNAi can help us to enlighten better about the gene functions in parasites, for targeted drug delivery against specific helminthes and finding the vaccine candidates, and reducing the role of vectors to transmit diseases. In this review, we provide the state of art information on RNAi phenomenon applied in the parasites, the prospects and possible pitfalls of this technique. Moreover, the factors required to obtain optimum results are discussed.
INTRODUCTION
RNA interference (RNAi) previously known as gene silencing is a technique of inhibition of gene expression in cells or organisms by introducing double–stranded RNA (dsRNA) leading to destruction of its homologous mRNA (Fire et al., 1998). It helps in analysis of gene function and has the potential in disease therapy, drug development and control of parasite transmission and development (Ullu et al., 2004). RNAi is a mechanism formerly believed to be defense machinery acting against the nucleic acids of external origin but it has been studied recently to have various other important mechanisms in protozoa like regulation of mRNA which gets accumulated in the cell before protein synthesis, surveillance of genome, rearrangement of DNA, silencing of chromatin as well (Nicolás et al., 2013). It appears promising in silencing gene expression in parasites by a process of specific target mRNA interference. Commonly used methods of delivery of dsRNA in protozoans and helminthes include soaking and electroporation where as in insect vectors with use of Micro–injection (Kang and Hong, 2008). However there are some sensible drawbacks when using RNAi in parasites viz., RNAi mechanism may have been eliminated in certain parasites, so identification of RNAi pathway in the targeted parasite is the prerequisite. A best delivery method should be established to generate optimal gene silencing and to reduce the non specific gene silencing. RNAi has been best studied in parasites like Trypanasoma brucei, Leishmania braziliensis (Atayde et al., 2013), Giardia lamblia, Trichinella spiralis (Chen et al., 2012), Entamoeba histolytica,(Zhang et al., 2011), Brugia malayi filarial parasite (Singh et al., 2012; Landmann et al., 2012), and Toxoplasma gondii (Barnes et al., 2012)
Discovery of RNAi
Gene silencing was first discovered in petunias plant (Napoli et al., 1990). A phenomenon called co suppression occurred due to high level of transgenic chalcone enzyme expression. This enzyme is responsible for normal colouration of the flowers but the transgenic flowers which showed over expression of this enzyme lost their normal pigmentation due to simultaneous suppression of mRNA of the transgene and endogenous chalcone enzyme (Caffrey, 2012). This phenomenon was further investigated by Craig Mello and Andrew Fire who clearly demonstrated the gene–silencing technique in Caenorhabditis elegans and called it as RNA interference in 1998 (Fire et al., 1998). For this discovery they were honored with Nobel Prize in Physiology or Medicine in 2006.
Components of RNAi
Double–stranded RNA (dsRNA) which may be of exogenous, viral or transposans dsRNA.
Dicer, a ribonuclease III enzyme which degrades dsRNA into 22 bp small interfering RNAs (siRNAs).
siRNA will induces the formation of RNA–induced silencing complex.
Argonaute (AGO), a group of proteins having three main domains viz., PAZ, MID and Piwi which helps in target identification and cleavage of the transcripts.
RNA–dependent RNA polymerase (RdRp) which triggers the synthesis of RNA from an RNA template.
Proteins like Systemic RNA interference–deficient (SID–1, SID–2) and RNAi spreading defective (RSD–4) which help in intake and spread of dsRNA in the cells.
RNAi Mechanism
RNAi is a kind of RNA–dependent gene silencing mechanism that is controlled by the RNA–induced silencing complex (RISC) and is initiated by short double–stranded RNA molecules which can from outside source or form the cell's own cytoplasm, where they interact with the catalytic RISC component argonaute. The double standard RNA transfected form outside into the cell takes a bit of journey into the cytoplasm where it is met with some of the enzymes which tame it before it reaches the destination of mRNA and acting on it to silence the gene. Double standard RNA once after its entry is spliced by the dicer into small ~20 bp length small RNA molecules with overhangs at both the ends. The source of dsRNA when it is from endogenous (originating in the cell), as in pre–microRNAs expressed from RNA–coding genes in the genome, miRNAs also follows the same sequence of events which the dsRNAs undergo like splicing by dicer and complexing with RISC. Thus, the two dsRNA pathways, exogenous and endogenous, converge at the RISC complex (Nykanen et al., 2001).
Exogenous or Endogenous dsRNA are the trigger material for the gene silencing mechanism. Dicer which is a ribonuclease III enzyme cleaves the long fragment of dsRNA into smaller fragments with a 2–nucleotide overhang at the 3' of both the ends. These short double–stranded fragments are called small interfering RNAs (siRNAs) (Elbashir et al., 2001). Till this step is the initial phase of RNAi and called as initiation process. This is followed by effector step where RISC binds with the small dsRNA. The small dsRNA are unwound into separate single stranded RNA and they are named as guide strand and passenger strand. During the course of the mechanism guide strand is bound to the RISC complex while the passenger strand is degraded. The guide strand then binds to the complementary strand of mRNA, which is ready for protein synthesis and this binding cause’s halt of protein synthesis. This binding also leads to the break in the mRNA sequence and these parts are degraded by enzymes (Wilson and Doudna, 2013). This whole mechanism leads to stop in protein synthesis and hence the gene, which has to be expressed, came to a halt and hence RNAi is called as gene silencing phenomenon (Tomari et al., 2004). The whole mechanism of RNAi is described in Figure 1.
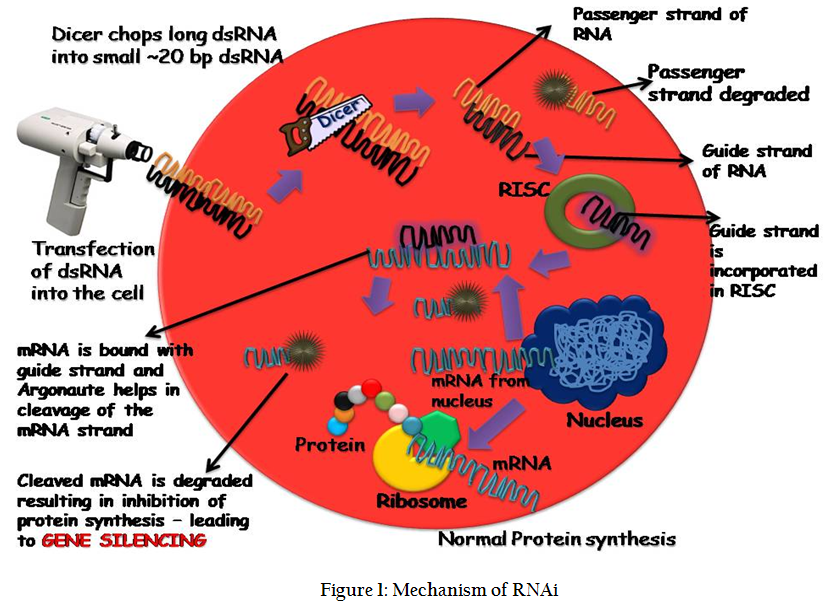
Methods of dsRNA Delivery
Mainly, four methods are used in parasites viz., i) feeding of Escherichia coli expressing double stranded RNA (dsRNA); (ii) soaking of short interfering (synthetic) RNA oligonucleotides (siRNA) or in vitro transcribed dsRNA molecules; and (iii) electroporation of siRNA or in vitro transcribed dsRNA molecules iv) Micro–injection dsRNA in to the organism.
RNAi in Helminths
Parasitic helminths can be controlled by use of RNAi technique, which will silence the critical genes necessary for normal physiological development of the worms. It can be applied in helminths to study the gene functions in the same way as it has been applied in other organisms. Genes identified using this technology can be used as drug and vaccine targets which is a promising therapeutic application.
At present only 10 species of animal parasitic helminth have been studied for RNAi effects since its successful application in C. elegans in 1998 (Geldhof et al., 2007). Animal parasitic helminthes, which are used for studying the RNAi, has been briefly described hereunder.
RNAi in Nematodes
RNA phenomenon was first reported in the animal nematode Nippostrongylus brasiliensis. Experiment was carried out in the nematode to suppress the expression of gene targeting acetylcholine esterase by using the RNAi technique. A 240 bp dsRNA was able to suppress the expression of AchE and it also silenced the effect of AchE for more than 6 days. By this way the expression of the AchE was decreased more than 90% which explained the effectiveness of this technique (Hussein et al., 2002).
An effective and promising method of control of Brugiya malai was developed using by targeting housekeeping genes like β–tubulin and RNA polymerase II large subunit. B. malayi is a mosquito transmitted filarial nematode and control of this nematode is not an easy job because of its transmission through mosquitoes. By advent of RNAi an effective control of this nematode was achieved by using 300 bp long dsRNA targeting the above mentioned targets (Aboobaker et al., 2003).
Next in the sequence of gene silencing was the Onchocerca volvulus L3 larvae by Lustigman and coworkers in 2004 who successfully reported gene silencing effect of RNAi. Cathespin L and cathespin Z like cysteine proteases was targeted during their study. These two cathespins were prerequisite for the nematode to molt form L3 stage larvae to L4 stage larvae. The method followed during this study was soaking L3 larvae in a solution containing dsRNA. There was a drastic reduction in the molting rate of 92% cathespin L and 86% for cathespsin Z which contributed significantly in the reduction of viability of L3 larvae by delaying the molting process for few days.
Subsequently the pig round worm Ascaris suum was used to study the RNAi effects by targeting inorganic pyrophosphate of the nematode which resulted in 31% reduction of molting of L3 to L4 larval stage (Islam et al., 2005). Though the inhibition of molting was lesser than that of O. volvulus, this paved a significant contribution to the RNAi phenomenon in Ascaris worms which lead to further exploitation of this useful technique in other related and distant round worms both in animals and humans.
RNAi experiment in sheep gastrointestinal nematode Trichostrongylus columbriformis suggested that siRNA and electroporation as more efficient molecules and also as a suitable delivery method to induce gene silencing by RNAi (Issa et al., 2005).Three different methods of delivery of dsRNA were tested like soaking, feeding and electroporation of siRNA. The targets used were ubiquitin and tropomyosin, as their DNA sequences are well conserved and readily available. The study concluded that tropomyosin gene and not ubiquitin gene was suppressed by feeding method when compared to other two methods whereas ubiquitin was reduced by both electroporation as well as by soaking method (Issa et al., 2005).
To get desired RNAi effects in Haemonchus contortus a voracious blood sucking nematode of sheep different conditions were tried targeting 11 different genes viz., β–tubulin, sec–23, Ca2+ binding protein, Heat Shock Protein (HSP70), vacuolar ATPase, cathepsin–L, paramyosin, Cu–Zn superoxide dismutase, intermediate filament, type IV collage and GATA transcription factor in different life stages L1 to L4 and adult parasite. The delivery methods used were feeding, soaking and electroporation. Optimum RNAi effects were seen in L3, L4 and adult stages of parasite where β–tubulin transcript levels were reduced when soaking method was used. Electroporation method failed to yield desired effects (Geldhof et al., 2006).
Similarly, RNAi effects in Ostertagia ostertagi a nematode parasite of cattle showed that the results of RNAi are not reproducible which indicated that RNAi delivery method requires further optimization to achieve optimal and consistent results (Visser et al., 2006).
Recently RNAi was applied in Trichenella spiralis, which is an intestinal nematode that infects more than 100 mammalian species, including humans. Gene silencing was used to determine the function of paramyosin which is a structural protein having an immunomodulatory properties. By suppressing paramyosin expression resulted in significant reduction in the parasite’s viability and infectivity, which confirmed role of paramyosin in the survival of T. spiralis and therefore it is considered as a promising vaccine candidate (Chen et al., 2012)
RNAi in Trematodes
The RNAi study in trematodes mainly emphasized on blood flukes called Schistosomes. In Schistosoma mansoni RNAi was used to know the function of cathepsin B an enzyme whose function was previously known to degrade host hemoglobin but with advent of RNAi it was proved that Cathepsin B was responsible for parasite growth, and not a mandatory for hemoglobin digestion (Correnti et al., 2005).
Similarly RNAi affects in developmental stages of schistosomes showing that RNAi has an affinity for particular developmental stage of the trematode. Boyle et al. (2003) targeted schistosome glucose transporter gene SGTP1, a facilitated diffusion glucose transporter and glyceraldehyde–3–phosphate dehydrogenase (GAPDH). They observed that RNAi affects in the form of reduced level of expression of both genes, which were transferable from miracidium, a free living larval stage to sporocyst stage in the life cycle of schistosomes. Interestingly, when dsRNAs were introduced at the sporocyst stage, the reduction was not observed (Tabara et al., 1998).
With the help of RNAi it was possible to assess the function of scavenger receptors of schistosomes, which are proved to be important in synthesis of biological membrane. When dsRNA targeting these receptors were delivered via, soaking method to miracidium stage of schistosomes, resulted in reduction of gene expression affecting normal morphology of sporocysts (Dinguirard et al., 2006).
In Schistosoma japonicum RNAi experiment targeting gynaecophoral canal protein which is responsible for correct sexual pairing resulted in 75% reduction of target gene transcripts so that pairing mechanism is altered and further inhibition of schistosomes development in the host.
Subsequently RNAi was applied to trematode like Fasciola hepatica to know the function of cathepsin L and B in parasitic invasion mechanism (Rinaldi et al., 2008).
Reports on RNAi experiment in cestodes is very scarce, recently robust knock–down of abundant gene transcripts was achieved using long dsRNA’s following short exposure times in Moniezia expansa, an intestinal tapeworm of sheep and goat (Pierson et al., 2010).
RNAi in Drug Target Discovery in Helminth Parasites
Discovery of drug targets in helminth parasites were tried using RNAi by a method called RNAi screen, which is used to screen abundant gene transcripts at a single time. This method met with desirable success in C. elegans but not with other helminthes. As this method required complex dsRNA delivery methods well characterized RNAi machinery which is present in C. elegans. With help of comparative genomics this can be applied in other nematodes for finding drug targets (Shuey et al., 2002). This approach has been tried in B. malayi where about 589 genes were identified which were critical for survival of this filarial nematode parasite (Jones et al., 2005).
Hurdles Ahead and Solutions to Sort out in the Field Of RNAi in Helminthes
RNAi is emerged as a successful technique only in C. elegans, but hurdle remains in other helminths where it showed either the technique is impossible or inconsistent to be adopted (Geldhof et al., 2007). This contrasting result may be due to difference in the delivery methods used and still there is no most efficient delivery method to introduce dsRNA in to the parasitic helminthes so as to get optimum RNAi effects. In some helminths absence of proteins like SID–1, SID–2, and RSD–4 which are involved in cellular intake and spread of dsRNA is an important factor for failure of RNAi (Peterson et al., 2007).
To achieve an uniform result among the pile of parasites, modification in the delivery methods has been suggested such as micro injection of dsRNA directly in to the pseudocoelom of helminthes there by bypassing barriers of dsRNA uptake in the gut lumen, making RNAi feasible in those heminths. Soaking and feeding methods failed to give desired effects (Viney and Thompson, 2008). Other modification suggested in the helminthes which are deficient in SID–1, SID–2, and RSD–4 proteins are heterologous expression of these proteins and generating RNAi effects (Winston et al., 2007).
Finally the question arises that why the other animal parasitic nematodes have different effect by RNAi while C. elegans shows promising results. It may be because of many reasons like different lifestyles of animal parasitic nematode. Possible risk of damage to nucleic acids may be greater for nematodes in animal hosts than it is for nematodes in plant hosts which may be related to differences in the ecology of these helminth parasites (Dalzell et al., 2012).
RNAi in Parasitic Protozoa
Protozoans that affect human being as well as animals include Plasmodium, Toxoplasma, Entamoeba, Giardia, Leishmania and Trypanosomes. Inspite of considerable impact of these protozoan parasites many aspects like their pathogenesis and their survivability within the host are still not completely understood. The discovery of RNAi has facilitated the gene function analysis in the protozoans which are not possible through conventional genetic approaches. Here we will focus on the current status of RNAi in studies of parasitic protozoa, with special emphasis on its use as a post genomic tool. (Kolev et al., 2011)
Trypanosomatids
Trypanosoma brucei was the first protozoan where RNAi was succeessfully applied. It has been used to study the functions of RNAi pathway proteins like Ago 1 and dicer as well as basic gene silencing mechanism. Using RNAi β– tublin gene expression in T. brucei was inhibited using tetracycline–inducible promoter (Ngo et al., 1998). Similarly RNAi effect was studied in T. congolense which causes Nagana disease in cattle. Here also similar results were obtained as that of T. brucei targeting β– tublin gene expression which resulted in changed morphology of transfected Trypanosomes. This suggested that RNAi machinery exists in T. congolense (Inoue et al., 2002).
Subsequent research in T. cruzi, Leishmania donovani and L. major suggested that RNAi pathway is absent in these protozoans as they lack Ago 1 protein which is essential for suppression of foreign and endogenous transgenes (DaRocha et al., 2004; Robinson and Beverley, 2003).
Apicomplexa
Protozoans belong to this group are having a specialized structure called apical complex that facilitates contact with the host cell. Plasmodium spp. protozoans fall under this apicomplexan phylum. RNAi effects were studied first in P. falciparum by using electroporation method dsRNA was delivered targeting dihydroorotate dehydrogenase (DHODH), an enzyme required for pyrimidine biosynthesis. Due to RNAi which resulted in reduction of DHODH levels causing a retardation of parasite growth. This proved RNAi machinery was present in P. falciparum. But subsequent studies showed that RNAi is still controversial and whether the RNAi pathway functions in Plasmodium or not (McRobert and McConkey, 2002).
RNAi research in Toxoplasma gondii suggests that it could be potentially present in Toxoplasma. A recent study shows that dsRNA targeting uracil phosphoribosyl transferase reduced the endogenous target transcript levels (Al–Anouti and Ananvoranich, 2002).
Other Protozoans
Database mining shows possibility of RNAi machinery in Giardia spp. of protozoans. Dicer protein was identified in Giardia intestinalis, which degraded dsRNA in to small RNAs. Recently RNase III and Ago2 proteins were characterized in Entamoeba histolytica which lead to the prediction that RNAi pathway is present in this group of enteric protozoan pathogens (Abed and Ankri, 2005).
RNAi pathway has been applied successfully in T. brucei using a vector system, which would generate dsRNAs under the control of an inducible promoter or with tissue–specific expression and this can be applied to other protozoan species. Primary perquisite is to identify RNAi machinery components should be identified and once it has been identified RNAi can be applied to study the gene functions to interfere parasite development for purpose of controlling them.
RNAi in Arthropods
Initially RNAi was successfully employed in model insect species like Drosophila melanogaster and in silkworm then subsequently applied to disease transmitting vectors like mosquitoes and ticks to study the gene functions. (Tuschl et al., 1999; Quan et al., 2002)
In Aedes aegypti which is the vector of dengue virus, RNAi was applied to induce resistance to viral infection in mosquito cells demonstrating application of RNAi in the control of vector borne diseases (Gaines et al., 1996).
Later RNAi pathway studied in Anophelese gambiae which is a main vector of malaria. By injecting dsRNA targeting an antimicrobacterial peptide gene called endogenous defensin gene which suppressed the defensin transcripts and thereby increasing the gram– positive bactiria activity in A. gambiae. This paved the way for possible application of of RNAi in elucidating gene functions in anopheline mosquitoe (Blandin et al., 2002)
Similarly gene silencing was applied identify 11 candidate immune genes which are called as antiplasmodial genes in A. gambiae using microarray analysis. RNAi was employed to analyze the functions of these genes by silencing of each immune gene which showed an increased Plasmodium levels (Dong et al., 2006).
Subsequently RNAi was used in Ixodes scapularis tick which is a vector of babesiois and lyme disease to analyse gene function. By injecting dsRNA against the Hazara virus (HAZV) gene inhibition of HAZV replication was achieved suggesting the application of RNAi in control of viral diseases and other infectious diseases in I. scapularis (Garcia et al., 2005).
Recently RNAi was employed to know the function of attacin gene in tsetse flies, Glossina spp which spread African trypanosomes like Trypanosoma brucei spp. experimental data suggests that the attacin is a refractory gene against African trypanosomes and also demonstrated that RNAi can be used as a powerful tool to investigate gene functions in tsetse flies (Nayduch and Aksoy, 2007).
In ticks RNAi is the most widely used method of genetic manipulations where use of other methods of genetic manipulations has been limited. It has been proved as a potential tool in studying tick gene function, characterization of the tick–pathogen interface and characterization of tick protective antigens (Kang and Hong, 2008)
Prospects of RNAi in Arthropods
Malaria control using germ line transformation and RNAi has been suggested. Mosquito genes such as leucine rich–repeat immune gene (LRIM1), C–type lectin (CTL4) and mannose binding CTL (CTLMA2) have been identified as candidates for malaria intervention since they are critical for parasite development. RNAi was used in tsetse flies to identify a refractory gene against African trypanosomes. With the identification of the tick genes that are responsible for pathogen development, RNAi can be employed to silence those genes and study their impact on pathogen development (Bell–Sakyi et al., 2007).
Further Concern about Optimization of RNAi in Parasites
The primary prerequisite is that RNAi pathway should be present in the targeted parasite.
Optimized culture conditions to maintain target parasite stage.
Measures for maximum uptake of dsRNA.
Whether RNAi effects are transferred from one stage to another stage in the life cycle of the parasite.
The target gene expression site
The target gene expression level
The capacity of RNAi to spread in the tissues where parasites are lodged.
The infectivity of RNAi – treated parasites to be examined in vivo (Knox et al., 2007)
CONCLUSIONS
Control of parasitic infection in the animal is the supreme need of the hour in developing country like India. There is a huge loss to the farmers due to parasitic infections and treatment aimed at them goes in vain leading to emergence of resistance in the parasite colony. Hence updated methods should be evolved so as to counteract these strategies and to minimize the parasitic load in animal. Search for vaccine is too long process and it has not yielded feasible results. RNA interference has the potential therapeutic and control applications in parasites. It helps in assessment of gene functions; identify vaccine candidates and drug targets. To get optimum RNAi effects in the parasites an effective delivery method is need of the hour. There are promising results in a handful of parasites and this list can be extended by following different delivery modes so as to get good transfection, and to allow the RNAi to do its job further. To achieve successful RNAi effects the mechanism of RNAi needs to be further characterized for each important parasites.
REFERENCES
Abed M and Ankri S (2005). Molecular characterization of Entamoeba histolytica RNase III and AGO2, two RNA interference hallmark proteins. Exp. Parasitol. 110: 265–269.
http://dx.doi.org/10.1016/j.exppara.2005.02.023
PMid:15955322
Aboobaker AA and Blaxter ML (2003). Use of RNA interference to investigate gene function in the human filarial nematode parasite Brugia malayi. Mol. Biochem. Parasitol. 129: 41–51.
http://dx.doi.org/10.1016/S0166-6851(03)00092-6
Al–Anouti F and Ananvoranich S (2002). Comparative analysis of antisense RNA, double–stranded RNA, and delta ribozyme–mediated gene regulation in Toxoplasma gondii. Antisense. and Nucleic Acid. Drug. Dev. 12: 275–281.
http://dx.doi.org/10.1089/108729002320351593
PMid:12238816
Atayde VD, Shi H, Franklin JB, Carriero N, Notton T, Lye LF, Owens K, Beverley SM, Tschudi C, Ullu E. (2013). The structure and repertoire of small interfering RNAs in Leishmania (Viannia) braziliensis reveal diversification in the trypanosomatid RNAi pathway. Mol Microbiol. 87(3): 580–93.
http://dx.doi.org/10.1111/mmi.12117
PMid:23217017 PMCid:PMC3556230
Barnes RL, Shi H, Kolev NG, Tschudi C, Ullu E. (2012). Comparative genomics reveals two novel RNAi factors in Trypanosoma brucei and provides insight into the core machinery. PLoS Pathog. 8(5): e1002678.
http://dx.doi.org/10.1371/journal.ppat.1002678
PMid:22654659 PMCid:PMC3359990
Bell–Sakyi L, Zweygarth E, Blouin EF, Gould EA and Jongejan F (2007). Tick cell lines: tools for tick and tick–borne disease research. Trends. Parasitol. 23: 450–457.
http://dx.doi.org/10.1016/j.pt.2007.07.009
PMid:17662657
Blandin S, Moita LF, Kocher T, Wilm M, Kafatos FC and Levashina EA (2002). Reverse genetics in the mosquito Anopheles gambiae: targeted disruption of the Defensin gene. EMBO Rep. 3: 852–856.
http://dx.doi.org/10.1093/embo-reports/kvf180
PMid:12189180 PMCid:PMC1084233
Boyle JP, Wu XJ, Shoemaker CB and Yoshino TP (2003). Using RNA interference to manipulate endogenous gene expression in Schistosoma mansoni sporocysts. Mol. Biochem. Parasitol. 128: 205–215.
http://dx.doi.org/10.1016/S0166-6851(03)00078-1
Caffrey CR (2012). RNA Interference: A Potential Discovery Tool for Therapeutic Targets of Parasitic Nematodes. Parasitic Helminths. DOI: 10.1002/9783527652969.ch6
http://dx.doi.org/10.1002/9783527652969.ch6
Chen X, Yang Y, Yang J, Zhang Z and Zhu X (2012). RNAi–Mediated Silencing of Paramyosin Expression in Trichinella spiralis Results in Impaired Viability of the Parasite. PLoS One 7(11): 49913.
http://dx.doi.org/10.1371/journal.pone.0049913
PMid:23185483 PMCid:PMC3503832
Correnti JM, Brindley PJ and Pearce EJ (2005). Long–term suppression of cathepsin B levels by RNA interference retards schistosome growth. Mol. Biochem. Parasitol. 143: 209–215.
http://dx.doi.org/10.1016/j.molbiopara.2005.06.007
PMid:16076506
Dalzell JJ, Warnock ND, Mcveigh P, Marks NJ, Mousley A, Atkinson L and Maule AG (2012). Considering RNAi experimental design in parasitic helminthes. Parasitol. 139 (5), 589–604.
http://dx.doi.org/10.1017/S0031182011001946
PMid:22216952
DaRocha WD, Otsu K, Teixeira SMR and Donelson JE (2004). Tests of cytoplasmic RNA interference (RNAi) and construction of a tetracycline–inducible T7 promoter system in Trypanosoma cruzi. Mol. Biochem. Parasitol. 133: 175–186.
http://dx.doi.org/10.1016/j.molbiopara.2003.10.005
PMid:14698430
Dinguirard N and Yoshino TP (2006). Potential role of a CD36–like class B scavenger receptor in the binding of modified low–density lipoprotein (acLDL) to the tegumental surface of Schistosoma mansoni sporocysts. Mol. Biochem. Parasitol. 146: 219–230.
http://dx.doi.org/10.1016/j.molbiopara.2005.12.010
PMid:16427708
Dong Y, Aguilar R, Xi Z, Warr E, Mongin E and Dimopoulos G (2006). Anopheles gambiae immune responses to human and rodent Plasmodium parasite species. PLoS Pathog. 2: 52.
http://dx.doi.org/10.1371/journal.ppat.0020052
PMid:16789837 PMCid:PMC1475661
Elbashir SM, Lendeckel W and Tuschl T (2001). RNA interference is mediated by 21– and 22–nucleotide RNAs. Genes. Dev. 15: 188–200.
http://dx.doi.org/10.1101/gad.862301
PMid:11157775 PMCid:PMC312613
Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE and Mello CC (1998). Potent and specific genetic interference by double–stranded RNA in Caenorhabditis elegans. Nature. 391: 806–811.
http://dx.doi.org/10.1038/35888
PMid:9486653
Gaines PJ, Olson KE, Higgs S, Powers AM, Beaty BJ and Blair CD (1996). Pathogen–derived resistance to dengue type 2 virus in mosquito cells by expression of the premembrane coding region of the viral genome. J. Virol. 70: 2132–2137.
PMid:8642634 PMCid:PMC190050
Garcia S, Billecocq A, Crance JM, Munderloh U, Garin D and Bouloy M (2005). Nairovirus RNA sequences expressed by a Semliki Forest virus replicon induce RNA interference in tick cells. J. Virol. 79: 8942–8947.
http://dx.doi.org/10.1128/JVI.79.14.8942-8947.2005
PMid:15994788 PMCid:PMC1168744
Geldhof P, Murray L, Couthier A, Gilleard JS, McLauchlan G, Knox DP and Britton C (2006). Testing the efficacy of RNA interference in Haemonchus contortus. Int. J. Parasitol. 36: 801–810.
http://dx.doi.org/10.1016/j.ijpara.2005.12.004
PMid:16469321
Geldhof P, Vissera A, Clarka D, Saundersa G, Brittona C, Gillearda J, Berrimana M and Knoxa D (2007). RNA interference in parasitic helminths: current situation, potential pitfalls and future prospects. Parasitol. 134: 609–619.
http://dx.doi.org/10.1017/S0031182006002071
PMid:17201997
Hoffmann KF (2004). An historical and genomic view of Schistosome conjugal biology with emphasis on sex–specific gene expression. Parasitol. 128: S11–S22.
http://dx.doi.org/10.1017/S0031182004006213
PMid:16454894
Hussein AS, Kichenin K and Selkirk ME (2002). Suppression of secreted acetylcholinesterase expression in Nippostrongylus brasiliensis by RNA interference. Mol Biochem. Parasitol. 122: 91–94.
http://dx.doi.org/10.1016/S0166-6851(02)00068-3
Inoue N, Otsu K, Ferraro DM, Donelson JE (2002). Tetracycline–regulated RNA interference in Trypanosoma congolense. Mol Biochem Parasitol; 120: 309–313.
http://dx.doi.org/10.1016/S0166-6851(02)00015-4
Islam MK, Miyoshi T, Yamada M and Tsuji N (2005). Pyrophosphatase of the roundworm Ascaris suum plays an essential role in the worm's molting and development. Infect. Immun. 73: 1995–2004.
http://dx.doi.org/10.1128/IAI.73.4.1995-2004.2005
PMid:15784540 PMCid:PMC1087427
Issa Z, Grant WN, Stasiuk S and Shoemaker CB (2005). Development of methods for RNA interference in the sheep gastrointestinal parasite, Trichostrongylus colubriformis. Int. J. Parasitol. 35: 935–940.
http://dx.doi.org/10.1016/j.ijpara.2005.06.001
PMid:16023650
Jones AK, Buckingham SD and Sattelle DB (2005). Chemistry–to–gene screens in Caenorhabditis elegans. Nat. Rev. Drug. Discov. 4: 321–330.
http://dx.doi.org/10.1038/nrd1692
PMid:15803195
Kang S and Hong YS (2008). RNA Interference in Infectious Tropical Diseases. Korean. J. Parasitol. 46(1): 1–15.
http://dx.doi.org/10.3347/kjp.2008.46.1.1
PMid:18344671 PMCid:PMC2526294
Knox DP, Geldhof P, Visser A and Britton C (2007). RNA interference in parasitic nematodes of animals: a reality check. Trends in parasitol. 23 (7): 105–107.
http://dx.doi.org/10.1016/j.pt.2007.01.007
PMid:17276139
Kolev NG, Tschudi C, and Ullu E (2011). RNA Interference in Protozoan Parasites Achievements and Challenges, Eukaryotic. Cell. 10(9):1156–1163
http://dx.doi.org/10.1128/EC.05114-11
PMid:21764910 PMCid:PMC3187059
Krautz–Peterson G, Radwanska M, Ndegwa D, Shoemaker CB and Skelly PJ (2007). Optimizing gene suppression in schistosomes using RNA interference. Mol. Biochem. Parasitol. 153: 194–202.
http://dx.doi.org/10.1016/j.molbiopara.2007.03.006
PMid:17420062
Landmann F, Foster JM, Slatko BK and Sullivan W (2012). Efficient in vitro RNA interference and immunofluorescence–based phenotype analysis in a human parasitic nematode, Brugia malayi. Parasit. Vectors. 5:16
http://dx.doi.org/10.1186/1756-3305-5-16
PMid:22243803 PMCid:PMC3292814
Lustigman S, Zhang J, Liu J, Oksov Y and Hashmi S (2004). RNA interference targeting cathepsin L and Z–like cysteine proteases of Onchocerca volvulus confirmed their essential function during L3 molting. Mol. Biochem. Parasitol. 138: 165–170.
http://dx.doi.org/10.1016/j.molbiopara.2004.08.003
PMid:15555728
McRobert L and McConkey GA (2002). RNA interference (RNAi) inhibits growth of Plasmodium falciparum. Mol. Biochem. Parasitol. 119: 273–278.
http://dx.doi.org/10.1016/S0166-6851(01)00429-7
Napoli C, Lemieux C and Jorgensen R (1990). Introduction of a chimeric chalcone synthase gene into petunia results in reversible co–sup–pression of homologous genes in trans. Plant Cell. 2: 291–299.
http://dx.doi.org/10.2307/3869076
http://dx.doi.org/10.1105/tpc.2.4.279
PMid:12354959 PMCid:PMC159885
Nayduch D and Aksoy S (2007). Refractoriness in tsetse flies (Diptera: Glossinidae) may be a matter of timing. J. Med. Entomol. 44: 660–665.
http://dx.doi.org/10.1603/0022-2585(2007)44[660:RITFDG]2.0.CO;2
Ngo H, Tschudi C, Gull K and Ullu E (1998). Double–stranded RNA induces mRNA degradation in Trypanosoma brucei. PNAS. 95: 14687–14692.
http://dx.doi.org/10.1073/pnas.95.25.14687
PMid:9843950 PMCid:PMC24510
Nicolás FE, Torres–Martínez S and Ruiz–Vázquez RM (2013). Loss and Retention of RNA Interference in Fungi and Parasites. PLoS Pathog. 9(1): 1003089.
http://dx.doi.org/10.1371/journal.ppat.1003089
PMid:23358725 PMCid:PMC3554617
Nykanen A, Haley B and Zamore PD (2001). ATP requirements and small interfering RNA structure in the RNA interference pathway. Cell. 107: 309–321.
http://dx.doi.org/10.1016/S0092-8674(01)00547-5
Pierson L, Mousley A, Devine L, Marks NJ, Day TA and Maule AG (2010). RNA interference in a cestode reveals specific silencing of selected highly expressed gene transcripts. Int. J. Parasitol. 40: 605–615
http://dx.doi.org/10.1016/j.ijpara.2009.10.012
PMid:19900457
Quan GX, Kanda T and Tamura T (2002). Induction of the white egg 3 mutant phenotype by injection of the double–stranded RNA of the silkworm white gene. Insect. Mol. Biol. 11: 217–222.
http://dx.doi.org/10.1046/j.1365-2583.2002.00328.x
PMid:12000640
Rinaldi G, Morales ME, Cancela M, Castillo E, Brindley PJ and Tort JF (2008). Development of functional genomic tools in trematodes: RNA interference and luciferase reporter gene activity in Fasciola hepatica. PLoS. Negl. Trop. Dis. 2, e260.
http://dx.doi.org/10.1371/journal.pntd.0000260
PMid:18612418 PMCid:PMC2440534
Robinson KA and Beverley SM (2003). Improvements in transfection efficiency and tests of RNA interference (RNAi) approaches in the protozoan parasite Leishmania. Mol Biochem. Parasitol. 128: 217–228.
http://dx.doi.org/10.1016/S0166-6851(03)00079-3
Shuey DJ, McCallus DE and Giordano T (2002). RNAi: gene–silencing in therapeutic intervention. Drug. Discov. Today. 7: 1040–1046.
http://dx.doi.org/10.1016/S1359-6446(02)02474-1
Singh M, Singh PK and Bhattacharya SM (2012). RNAi mediated silencing of ATPase RNA helicase gene in adult filarial parasite Brugia malayi impairs in vitro microfilaria release and adult parasite viability. J. Biotechnol. 157(3): 351–358.
http://dx.doi.org/10.1016/j.jbiotec.2011.12.003
PMid:22192512
Tabara H, Grishok A and Mello CC (1998). Reverse genetics: RNAi in C. elegans: soaking in the genome sequence. Science. 282: 430–431.
http://dx.doi.org/10.1126/science.282.5388.430
PMid:9841401
Tomari Y, Du T, Haley B, Schwarz DS, Bennett R, Cook HA, Koppetsch BS, Theurkauf WE and Zamore PD (2004). RISC assembly defects in the Drosophila RNAi mutant armitage. Cell. 116: 831–841.
http://dx.doi.org/10.1016/S0092-8674(04)00218-1
Tuschl T, Zamore PD, Lehmann R, Bartel DP and Sharp PA (1999). Targeted mRNA degradation by double–stranded RNA in vitro. Genes. Dev. 13: 3191–3197.
http://dx.doi.org/10.1101/gad.13.24.3191
PMid:10617568 PMCid:PMC317199
Ullu E, Tschudi C and Chakraborty T (2004). RNA interference in protozoan parasites. Cell. Microbiol. 6: 509–519
http://dx.doi.org/10.1111/j.1462-5822.2004.00399.x
PMid:15104593
Ullu E, Lujan HD and Tschudi C (2005). Small sense and antisense RNAs derived from a telomeric retroposon family in Giardia intestinalis. Eukaryot Cell. 4: 1155–1157.
http://dx.doi.org/10.1128/EC.4.6.1155-1157.2005
PMid:15947207 PMCid:PMC1151983
Viney ME and Thompson FJ (2008). Two hypotheses to explain why RNA interference does not work in animal parasitic nematodes. Int. J. Parasitol. 38: 43–47.
http://dx.doi.org/10.1016/j.ijpara.2007.08.006
PMid:18028931
Visser A, Geldhof P, de Maere V, Knox DP, Vercruysse J and Claerebout E (2006). Efficacy and specificity of RNA interference in larval lifestages of Ostertagia ostertagi. Parasitol. 31: 1–7.
Wilson RC and Doudna JA (2013). Molecular Mechanisms of RNA Interference. Annual Annu. Rev. Biophys. 42: 217–239
http://dx.doi.org/10.1146/annurev-biophys-083012-130404
PMid:23654304
Winston WM, Sutherlin M, Wright AJ, Feinberg EH and Hunter CP (2007). Caenorhabditis elegans SID–2 is required for environmental RNA interference. PNAS. 104: 10565–10570.
http://dx.doi.org/10.1073/pnas.0611282104
PMid:17563372 PMCid:PMC1965553
Zhang H, Pompey JM and Singh U (2011). RNA interference in Entamoeba histolytica: implications for parasite biology and gene silencing. Future Microbiol. 6:103–117.
http://dx.doi.org/10.2217/fmb.10.154
PMid:21162639 PMCid:PMC3038252

