
Advances in Animal and Veterinary Sciences


Review Article
Advances in Animal and Veterinary Sciences 2 (4S): 33 – 41Special Issue – 4 (2014) (Reviews on Frontiers in Animal and Veterinary Sciences)
Evolution and Spread of Avian Influenza H5N1 Viruses
Chakradhar Tosh*, Shanmugasundaram Nagarajan, Harshad V. Murugkar, Sandeep Bhatia, Diwakar D. Kulkarni
-
High Security Animal Disease Laboratory, Indian Veterinary Research Institute, Bhopal – 462 021, India
*Corresponding author:[email protected]
ARTICLE CITATION:
Tosh C, Nagarajan S, Murugkar HV, Bhatia S, Kulkarni DD (2014). Evolution and spread of avian influenza H5N1 viruses. Adv. Anim. Vet. Sci. 2 (4S): 33 – 41.
Received: 2014–02–26, Revised: 2014–05–03, Accepted: 2014–05–05
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/journal.aavs/2014/2.4s.33.41
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
The Asian lineage of H5N1 avian influenza virus emerged from geese (A/Goose/Guangdong/1/96) in China in 1996, and is continuing its spread infecting domestic poultry, other birds and sporadic infection to humans revealing its pandemic potential. Since late 2003, H5N1 virus has been reported over 60 counties in Asia, Europe and Africa killing millions of poultry and infected humans in 16 countries with a total of 650 confirmed cases including 386 fatal have been reported to WHO. Spread of H5N1 virus has been linked with bird migration, although trade of poultry and poultry products has also been attributed in the spread. During the last one and half decades of circulation in poultry, the H5N1 virus has undergone significant genetic diversification leading to emergence of 10 major virus clades (clades 0-9), with subsequent diversification into 2nd-, 3rd- and 4th- order clades. Vaccination is one of the strategies for control and prevention of H5N1 virus infection; however, some countries still face challenges to eliminate H5N1 virus in poultry. One of the major problems is antigenic drift in viruses that develops due to vaccine pressure, particularly when vaccination is not implemented properly. Two classes of anti-viral drugs (neuraminidase inhibitors and adamantanes) are available for treatment and prophylaxis of influenza; however, there is emergence of drug resistant variants in poultry and humans. Wide-spread circulation and ever changing nature of H5N1 virus make avian influenza control program more difficult. Therefore, educational programs to enhance awareness on avian influenza, animal husbandry practices and biosecurity measures are important in control of the disease.
INTRODUCTION
Avian influenza virus, particularly the Highly Pathogenic Avian Influenza (HPAI) virus subtype H5N1, is evolving and continuing its spread. The ancestor of the currently circulating H5N1 virus was emerged from geese (A / Goose / Guangdong / 1 / 96, Gs / GD / 96) in southern China (Guangdong province), in 1996 (Xu et al., 1999). Subsequently, the H5N1 virus was detected during outbreaks in chickens in Hong Kong in April 1997 before jumping to humans (Suarez et al., 1998; Subbarao et al., 1998). In May 1997, a child (3 – year – old boy) was found positive for H5N1 virus who subsequently died of respiratory failure on 21st of May (Subbarao et al., 1998). This is the first report of H5N1 virus (A / Hong Kong / 156 / 97, HK / 97) infecting humans. By the end of 1997, the H5N1 virus had infected 17 more people in Hong Kong. A total of six deaths were confirmed ( CDC 1998). Destruction of all poultry in Hong Kong eliminated the deadly H5N1 virus (Guan et al., 2002b). However, importation of ducks and geese led to the introduction of Gs / GD / 96 – like virus into Hong Kong which subsequently reassorted with other subtypes to generate different reassortants (Guan et al., 2002a, 2002b). In late 2002, the first reported incidence of the spread of H5N1 virus from domestic to wild aquatic birds occurred in two parks in Hong Kong (Ellis et al., 2004). Between late 2003 and early 2005, H5N1 HPAI viruses spread to other regions of East and Southeast Asia leading to death or slaughter of 150 million poultry (Chen et al., 2006a). In late April 2005, a large outbreak of H5N1 virus infection occurred in different species of migratory waterfowls at Qinghai Lake in central China leading to death of more than 6000 wild birds (Chen et al., 2005). This is the first instance of H5N1 HPAI virus causing such heavy mortality in wild birds. Four different H5N1 genotypes were detected during the outbreak; however, one of these became dominant and spread westward and by the end of 2005, the H5N1 virus had spread to Central Asia and Europe. In early 2006, the virus spread to other European countries infecting poultry and wild birds. The virus spread to South Asia and Africa with report of H5N1 virus infection in poultry in India, Pakistan and Nigeria during February, 2006. The H5N1 virus has been detected over 60 countries across Asia, Europe and Africa killing domestic poultry, other birds and occasional transmission to humans thereby raising the pandemic potential of the virus.
ETIOLOGY AND EVOLUTIONARY STRATEGIES
The H5N1 avian influenza virus (AIV) is a member of the genus Influenzavirus A (type species: Influenza A virus) under Orthomyxoviridae family (Forrest and Webster, 2010). The viral genome is comprised of eight RNA segments of negative polarity that encodes at least for 11 distinct polypeptides. The polypeptides are three surface proteins [HA, NA and matrix 2 (M2)], three polymerases [polymerase basic (PB) 1, PB2 and polymerase acidic (PA)], PB1-F2, nucleoprotein (NP), matrix 1 (M1), non-structural protein (NS1) and nuclear export protein (NEP). Influenza A viruses (IAVs), like other RNA viruses, undergo rapid changes due to its error-prone replication of the polymerase complex leading to accumulation of point mutations resulting in antigenic changes (commonly known as “antigenic drift” ). antigenic drift is a gradual changes in the nucleotide sequence and is detected in all genome segments of the virus; however it is more evident within the genes that code for antibody-binding sites such as the HA and NA. antigenic drift is shaped by selection pressure acting along the entire genome of virus. antigenic drift is responsible for annual influenza epidemics in humans. The segmented genome of IAV gives an added advantage of exchanging gene segments between viruses (known as “antigenic shift”) when more than one genotype of the virus infects a single host cell leading to emergence of a novel virus. Historically, antigenic shift is responsible for emergence of pandemic strains to which human population is naive.
GENOME DIVERSITY
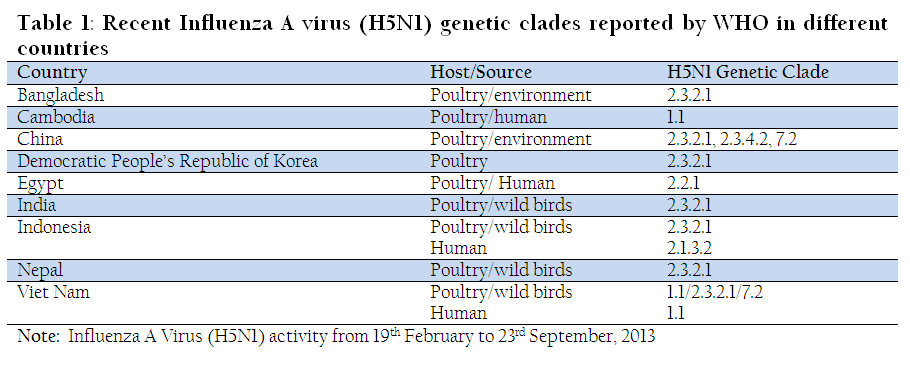
Since 1959, over 20 HPAI outbreaks have been recorded in poultry, of which three (first one in chickens in Scotland, 1959; second in turkey in England, 1991 and the third started in late 2003 and is continuing in poultry and other birds) were caused by H5N1 subtype (Alexander et al., 2007). Out of the above three, only the descendants of Gs / GD / 96 -like viruses have spread a large geographical area and persisted for such a long period of time (still continuing). The H5N1 HPAI virus (Gs / GD / 96) that emerged from geese in 1996 in China is evolving, and over the last one and half decades of circulation infecting multiple hosts including domestic poultry, wild birds, cats, dogs, tigers and mammals led to evolution of distinct genetic lineages. Publications describing these results used different nomenclatures to classify same groups of viruses within the Gs / GD / 96 – like lineage (Chen et al., 2006b; Salzberg et al., 2007) leading to confusion in the interpretation of results. To address these issues, an international core group of scientists were engaged to formulate a uniform nomenclature of the H5N1 viruses using phylogenetic analysis of the HA gene (WHO / OIE / FAO H5N1 Evolution Working Group, 2008). Initial report identified ten distinct clades (clades 0 – 9) of H5N1 Influenza A viruses that emerged from Gs / GD / 96 – like lineage (Figure 1). The clades are defined on the basis of phylogenetic analyses and nucleotide sequence divergence of a monophyletic group with ≥60 % bootstrap support at the clade-defining node. The intra – and inter-clade sequences maintain an average percentage divergence of ≤ 1.5% and > 1.5%, respectively. It was observed that certain clades became dominant and had wide geographical distribution. For example, clade 2 and its second order clade 2.2 had a wide geographical distribution in Asia, Europe and Africa. The other major clades identified were clade 0, 2.1, 2.3, 2.4 and 2.5. Subsequent analysis with contemporary H5N1 virus sequences reveals that a number of clades including clades 0, 3, 4, 5, 6, 8 and 9 and some second and third order groups of the clade 2 had not been detected during 2008 or before (WHO / OIE / FAO H5N1 Evolution Working Group, 2012). However, other H5N1 clades that continue to circulate in different regions are clades 1 (Cambodia and Vietnam), 2. 1. 3 (Indonesia), 2. 2. 1 (Egypt), 2. 2. 2 (India, Bangladesh and Bhutan), 2. 3. 2 (China, Hong Kong, Japan, Korea and Vietnam) 2. 3. 4 (China, Hong Kong, Myanmar, Vietnam), and 7 (Vietnam). Among the circulating clades, clade 2. 3. 2 and its subsequent divergent clade 2.3.2.1 (4th order clade) have wide geographical spread in Asia including China, Hong Kong, Korea, Vietnam, Laos, Bangladesh, Bhutan, India, Indonesia, Nepal, Myanmar, Iran, Mongolia and Russia, and in Europe including Bulgaria and Romania. Certain clades were restricted to specific location such as clade 1 (Cambodia, Vietnam and Malaysia), clade 2. 1. 3 (Indonesia), clade 2. 2. 2 (Bangladesh, Bhutan and India), clade 2 .2. 1 (Egypt and Israel), clade 2. 3. 4 (China, Hong Kong, Vietnam, Thailand, Myanmar and Laos), and clade 7 (China and Vietnam).
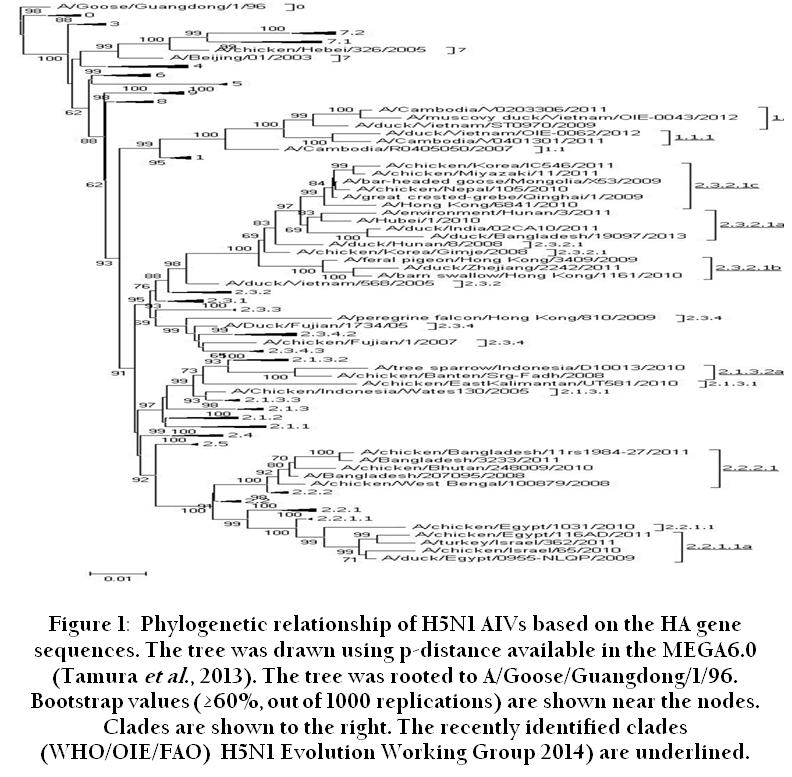
Antigenic diversity between the clades complicates the control of H5N1 infection through vaccination, as clades generally do not cross neutralize. Very recently, eight new clades have been recommended, based on new H5N1 isolates collected during 2011 and 2012, in clade 1. 1 (Vietnam), 2. 1. 3. 2 (Indonesia), 2. 2. 2 (India / Bangladesh), 2. 2. 1. 1 (Egypt / Israel), and 2. 3. 2. 1 (Asia) ( WHO / OIE / FAO H5N1 Evolution Working Group 2014). Since the H5N1 viruses are continuing to evolve rapidly, the WHO is coordinating the selection of representative candidate vaccine viruses as a pre-pandemic preparedness. Recently circulating H5N1 clades in different geographical locations reported by WHO are given in Table 1.
REASSORTMENT OF VIRUSES
Reassortment is a natural process in which viruses possessing segmented genomes exchange gene segments among themselves. Inter-subtype reassortment is one of the strategies of the H5N1 HPAI virus evolution. Detection of reassortment and subsequent genotyping of virus is carried out by phylogenetic analysis of all the eight gene segments of the virus. Reassortment supports rapid growth of the variant virus compared to the parent viruses. Analysis of all eight-gene segments has identified that Gs / GD / 96 virus has undergone extensive genetic reassortments with different IAVs leading to generation of several distinct genotypes. The HK / 97 virus, a known avian virus, that caused severe human infection and death is a double / triple reassortant with the HA gene from Gs / GD / 96-like virus, the NA and the six internal genes from H6 subtype or H9N2 virus found in terrestrial poultry (Guan et al., 2002a; Cheung et al., 2007).
The Hong Kong 1997 virus was not detected after destruction of all poultry in Hong Kong area. However, the Gs / GD / 96-like virus had undergone extensive genetic reassortment and produced number of genotypes (genotypes A, B, C, D and E) that were detected in poultry in markets in Hong Kong during 2001 (Guan et al., 2002b). A new reassortant virus (genotype Z) evolved in 2002 and spread to other Southeast Asian countries including Southern China, Cambodia, Indonesia, Thailand and Vietnam that caused widespread outbreaks in chickens and ducks during 2003 and 2004. In the same time, another new genotype V was detected in Japan and South Korea (Li et al., 2004). In late April 2005, four genotypes were detected from dead wild birds in Qinghai lake, however, the dominant genotype Z subsequently spread to Russia, Europe, Africa and other Asian countries including India, Pakistan, Afghanistan, Israel etc. (Chen et al., 2006a). In 2005, the “Fujian – like” lineage (clade 2.3.4) emerged from poultry in China and spread to Laos, Malaysia and Thailand (Smith et al., 2006), which was subsequently renamed as genotype V (Duan et al., 2008). Comprehensive sequence analysis of H5N1 viruses isolated during 1996 to 2006 in China detected 44 reassortants / genotypes including 34 transient and 10 persistent, except HK/97 virus (Duan et al., 2008). The novel “Hubei – like” virus was detected in Hubei province of China in 1997, which was a reassortant with internal genes are closely related to viruses isolated during 1970s from aquatic birds. A novel genotype P was detected in Laos in 2007 which was a reassortant between the Gs/GD /96-like virus with the PB1 and PB2 genes closely related to the Eurasian aquatic viruses (Boltz et al., 2010). Subsequently, reassortments were detected in H5N1 viruses in other regions including Bangladesh, Nigeria, Russia and Vietnam (Lipatov et al., 2007; Fusaro et al., 2010; Tran et al., 2011; Monne et al., 2013; Gerloff et al., 2014).
AVIAN-TO-HUMAN TRANSMISSION
Avian influenza is primarily a disease of birds. However, it has been seen, although sporadic, to cross the species barriers to infect humans. The first cases of human infection with avian influenza virus (subtype H5N1) were reported in Hong Kong during 1997, where eighteen individuals were infected leading to six deaths. Subsequently, two people in Hong Kong found positive for H5N1 virus infection leading to one death during 2003 (Peiris et al., 2004). Since late 2003, H5N1 virus has been reported infecting humans, and as of 24th January 2014 a total of 650 confirmed cases of which 386 fatal have been reported to WHO from 16 countries around the world (www.who.int). The Influenza A virus infection occurs through binding of viral HA and the oligosaccharide (sialic acid) molecule of the host cell. The avian and human Influenza A viruses differs in their recognition to host cell receptors. The HA from avian strains preferentially bind to sialic acids connected to sugar (galactose) by an α2, 3 linkage (SA α 2, 3 Gal), whereas the HA from human strains preferentially bind to sialic acids connected to galactose by an α2, 6 linkang (S A α 2 ,6 Gal) ( Imai and Kawaoka, 2012). The receptor recognition shift is one of the changes that must occur for efficient transmission of H5N1 virus in humans for pandemic spread. Pigs are considered as intermediate host or “mixing vessels” for the reassortment to occur between the human and avian influenza viruses as the tracheal epithelial cells express both α 2 , 3 and α 2 , 6 linked sialic acids. However, direct transmission of H5N1 virus from chickens to humans that occurred in 1997 in Hong Kong demonstrated for the first time that receptor specificity of avian influenza viruses may not restrict initial avian-to-human transmission (Matrosovich et al., 1999). The characteristic features of the 1997 Hong Kong chicken H5N1 viruses had a deletion (19 – amino – acids) in the NA stalk region and the globular head of the HA possessed a carbohydrate molecule compared to H5 viruses detected in wild aquatic birds. This finding suggested that alterations in both the surface glycoproteins HA and NA of the virus is required for adaptation of AIVs from wild aquatic birds to domestic poultry (Matrosovich et al., 1999). Amino acid mutations Gln222Leu and G l y 2 2 4 S e r (H5 numbering) in HA alter switching of IAVs binding from avian – like receptor to human-like receptor. A recent study demonstrated that a mutant virus combining G l n 1 9 6 A r g with G l n 2 2 2 L e u and G l y 2 2 4 S e r resulted in the shift from avian-like to human-like receptor switching (Chen et al., 2012). Table 2 shows the amino acids present in the receptor binding sites of H5N1 viruses isolated in different host species.

WILD MIGRATORY BIRDS – THEIR ROLE IN THE SPREAD AND EVOLUTION OF H5N1 VIRUS
In December 2002, mortality in wild and captive birds was reported due to H5N1 virus infection at Penfold and Kowloon Parks in Hong Kong (Ellis et al., 2004). This lethality of H5N1 virus to aquatic birds is first incidence since 1961, when Common Terns were killed by H5N3 virus in South Africa (Becker et al., 1966). In late April 2005, an outbreak of H5N1 virus infection occurred in Qinghai Lake (China) in which massive death of wild migratory birds was reported and the dominant virus of clade 2. 2 was detected from dead birds (Chen et al., 2005; 2006a). Subsequently, clade 2. 2 virus spread from Asia to Europe and Africa and the role of wild migratory birds in the dissemination of the H5N1 virus has been highlighted (Chen et al., 2006b; Kilpatrick et al., 2006; Fusaro et al., 2010). The H5N1 virus has been detected in several countries from wild birds predominantly in swans (whooper swan and mute swan), and a small number of cases in other species such as gulls, herons and raptors (Ellis et al., 2004; Teifke et al., 2007; Starick et al., 2008; Bröjer et al., 2009). By integrating genetic data, bird migration and, trade of poultry and wild birds, it was demonstrated that spread of H5N1 virus to most countries (i.e., 20 out of 23) in Europe was most likely through migratory bird compared to introduction in Asia (9 out of 21 introductions) and Africa (3 out of 8 introductions) (Kilpatrick et al., 2006). The H5N1 virus (clade 2. 2) isolated from Lake Qinghai was dominant during 2005 – 2007, whereas the new clade 2. 3. 2 overtook in 2008 and continuing its spread and diverged into 4th order clade 2. 3. 2. 1 (Smith et al., 2009; Reid et al., 2011; Nagarajan et al., 2012; Khan et al., 2013). The dominance of clade 2. 3. 2 and its subsequent divergent clade 2. 3. 2. 1 could partly be due to the presence of the virus in wild birds and continuous circulation in the H5N1 endemic regions. Recent reports revealed further genetic diversity in clade 2. 3. 2. 1 viruses in Asia (Creanga et al., 2013; WHO 2014). The H5N1 viruses from clades 2. 3. 2 and 2. 3. 4 were detected during 2004-2008 from dead wild birds such as egrets, falcons, herons and various passerine birds in Hong Kong (Smith et al., 2009). Subsequently, clade 2. 3. 2 virus was detected in poultry and wild birds during 2008 – 2009 in Japan, Korea, Russia, Mongolia and China (Kang et al., 2011; Sharshov et al., 2010; Li et al., 2011), and by 2010 / 2011 it was spread to Bulgaria and Romania in Europe and Bangladesh and India in South Asia (Reid et al., 2011; Nagarajan et al., 2012). Clade 2.3.4 virus (“Fujian – like”) that emerged during 2005 in poultry in China, subsequently spread to Hong Kong, Vietnam, Thailand, Myanmar, Laos and Bangladesh (Smith et al., 2006; Pfeiffer et al., 2011). Other minor clades isolated from wild birds were clades 1. 2. 2, 2. 5, 6, and 7 (Starick et al., 2008; Kou et al., 2009; Xiao et al., 2013) indicating genetic diversity of H5N1 viruses in wild birds and posing threat for wide spread.
EMERGENCE OF DRUG-RESISTANCE
Currently two classes of anti-viral drugs targeting the NA and M2 proteins are being used for treatment or prophylaxis of influenza. The NA inhibitors such as oseltamivir and zanamivir bind to the enzyme active site preventing the release of progeny virus from host cells (Hurt et al., 2009).
The M2 inhibitors such as amantadine and rimantadine block the viral M2 ion channel activity of virus, thereby preventing viral entry into host cells. Unfortunately, there is emergence of viral resistance to both the classes of drugs. Mutations in the drug binding sites are the most important cause of viral resistance, primarily due to the error-prone replication of the viral polymerase complex. A study highlights that H5N1 drug resistance, in most cases, is due to point mutations rather than genetic reassortment (Hill et al., 2009). Further, they also found that some M2 positions that confer resistance to adamantane derivatives are under positive selection, but no positive selection was found on NA that confer resistance to oseltamivir. Mutations at drug binding sites (G l u 1 1 9 V a l, H i s 2 7 4 T y r, A r g 2 9 2 L y s and A s n 2 9 4 S e r, N2 numbering) in the NA could confer viral resistance to drug oseltamivir (Govorkova et al., 2013) with substitutions at positions 274 and 294 are most common. Resistance to drug oseltamivir with an alteration H i s 2 7 4 T y r in NA in H5N1 viruses have been reported in patients with oseltamivir treatment or prophylaxis (Le et al., 2005). Another study reported reduced (57 to 138 – fold) susceptibility of H5N1 viruses to oseltamivir in patients in Egypt associated with Asn294Ser (A s n 2 9 5 S e r according to N1 numbering) mutation in the NA (Earhart et al., 2009). Besides involvement of the NA positions at 274 and 294, amino acid mutations V a l 1 1 6 A l a, I l e 1 2 2 L e u, L y s 1 5 0 A s n and S e r 2 4 6 A s n associated with reduced susceptibility to oseltamivir have been reported in Laos during 2006-2008 with clade 2. 3. 2 viruses (Boltz et al., 2010). Analysis of global sequence data of H5N1 viruses isolated during 2002 – 2012 revealed that the markers for NA inhibitor resistance (G l u 1 1 9 A l a, H i s 2 7 4 T y r and A s n 2 9 4 S e r) were less (0.8%) in avian isolates compared to human (2.4%) isolates, whereas the marker of reduced susceptibility (I l e 1 1 7 V a l, L y s 1 5 0 A s n, I l e 1 2 2 V a l / T h r / L y s and S e r 2 4 6 A s n) were more (2.9%) in avian isolates compared to human (0.8%) isolates (Govorkova et al., 2013). A Glu119Ala substitution associated with resistance of H5N1 viruses to drug zanamivir has been reported.
Molecular changes in the transmembrane region of the M2 ion channel protein of influenza viruses are responsible for the emergence of amantadine and rimantadine drug-resistance. Single or multiple amino-acid mutations have been identified at residues L e u 2 6, V a l 2 7, A l a 3 0, S e r 3 1, and G l y 3 4, with mutations at position 31 of M2 are the most common (Cheung et al., 2006; Govorkova et al., 2013). Amantadine-resistance with Ser31Asn mutation in M2 protein of H5N1 viruses have been reported in poultry during 2001-2005 in Northern China (Hebei province) where the drug was being used extensively in poultry farms (He et al., 2008). Amantadine-resistant mutants in H5N1 virus have been reported from Hong Kong SAR, mainland China, Vietnam, Thailand, Indonesia, Cambodia, Malaysia, Saudi Arabia and India with Ser31Asn substitutions in majorities of the cases (Cheung et al., 2006; Hurt et al., 2007; Monne et al, 2008; Tosh et al., 2011a). The frequency of amantadine-resistant H5N1 viruses detected during 2002-2012 was lower among avian (31.6%) than human (62.2%) viruses and are disproportionately distributed through different HA clades (Govorkova et al., 2013). It has been observed that double (L e u 2 6 I l e and S e r 3 1 A s n) mutations in M2 protein are more often than a single (S e r 3 1 A s n) mutation in both avian and human isolates. Analysis in the in vitro system found that the above double mutations in M2 protein conferred a growth advantage to the H5N1 virus, especially at 400C, which may contribute to the efficient replication of the virus in the intestine of aquatic birds (Cheung et al., 2010). In recent years (2010-2012), an increase in amantadine-resistant in H5N1 viruses has been observed in avian species, and is more likely due to drug selection pressure (Govorkova et al., 2013). This conclusion is supported by field data on use of amantadine in poultry in Northern China (He et al., 2008).
EMERGENCE OF VARIANTS IN VACCINATED POULTRY
To minimize the direct loss caused by H5N1 HPAI viruses, vaccination of poultry has been used extensively as a tool for control and prevention in many endemically infected countries including China, Egypt, Indonesia and Vietnam (Domenech et al., 2009). However, the view of the FAO on vaccination is a legitimate aid and not as a panacea in control and prevention HPAI virus infection. Despite the successes with vaccination strategies in reducing infection in poultry and human, some countries still face challenges to eliminate H5N1 virus in poultry (Domenech et al., 2009). One of the major problems is drift in antigenicity in viruses that develops due to vaccine pressure, particularly when vaccination is not implemented properly (Savill et al., 2006; Wang et al., 2012). Antigenic variants have been detected in China, Egypt and Indonesia following H5N1 vaccination (Domenech et al., 2009; Cattoli et al., 2011; Wang et al., 2012). A complex genetic drift in HA has been reported in Egyptian H5N1 viruses under AI vaccine pressure where high mutations were observed at residues 43 and 71 (epitope E), 110 (epitope A), 129 (epitope A and receptor binding site), 140, 141 and 144 (epitope B), 151, 185, 192 and 226 (epitope D) and 154 (epitope D and glycosylation site) (Abdel-Moneim et al., 2011). Another study with H5N1 viruses isolated during 2006-2012 in Egypt highlighted 28 amino acid changes in the HA antigenic sites, which could be due to selection pressure from vaccination and / or natural infection (Ibrahim et al., 2013). The HA amino acid substitution has led to significant antigenic drift (30. 7 – 79.1% antigenic relationship) of the recent isolates compared to the ancestral viruses. Therefore, it is possible that the mass vaccination in China, Egypt and Indonesia might have played a role in the emergence of antigenic drift in currently circulating H5N1 viruses. Recent in vitro studies with H5N1 escape mutants supports the hypothesis that the amino acid changes in the HA antigenic epitopes are due to vaccine pressure (Hoper et al., 2012; Sitaras et al., 2014).
EVOLUTION RATE AND POPULATION DYNAMICS
IAVs, like other RNA viruses, undergo error-prone replication in their genome due to lack of proof reading activity of RNA polymerase complex (Forrest and Webster, 2010). The high error rate leads to generation and amplification of viruses with similar but non-identical genomes, a phenomenon called “quassispecies”, in which each individual genotype has different fitness for the host (Domingo et al., 1985). Further this high error replication is an advantageous phenomenon for the IAVs to replicate in the new environment, especially when switching of host occurs (Suarez, et al., 2000). The rate of evolution, which is calculated from nucleotide substitutions (synonymous and non-synonymous changes), can be determined from sequences of a monophyletic cluster sampled over a period of time. The evolutionary rates calculated for surface genes of H5N1 viruses (H5 – 4.77×10−3 substitutions / site / year and N1 – 5.19×10−3 substitutions / site / year) of Gs / GD / 96-lineage are higher than that of internal genes (1.84–2.62×10−3 substitutions / site / year) (Vijaykrishna et al., 2008). However, the rate of evolution of H5N1 clade 2 . 2 . 1 viruses isolated from Egypt during February 2006 to July 2010 was marginally higher (5.36 x 10-3 substitution / site / year), and could be attributed to mass vaccination campaign in poultry (Cattoli et al., 2011). Within clade 2. 2. 1, the viruses were further divided into two major subclades (2. 2. 1A and 2. 2. 1B), and the rate of evolution within the subclades was markedly different. A mean rate of evolution was estimated to be 4.07 x 10-3 substitution / site / year for subclade 2.2.1A and 8.87 x 10-3 substitution / site / year for subclade 2. 2. 1B. The difference in mean evolutionary rates between two subclades coupled with stronger signature of positive selection in viruses circulating in Egypt since 2007 indicates vaccine selection. Significant increase in substitution rates in HA1 gene has been observed during period of mass vaccination (2005 – 2010 in China and 2003 – 2009 in Indonesia), in contrast to the periods when no vaccination program was undertaken (1996 – 2004 in China and 2004 – 2008 in Thailand) (Wang et al., 2012).
Using latest bioinformatics tools such as dated phylogenies, the time of most recent common ancestors (TMRCA) of each gene segments of a virus is identified. Based on the TMRCA, it was proposed that the Gs / GD / 96 virus was introduced into poultry from wild birds in January 1994 (95% confidence) (Vijaykrishna et al., 2008) as low pathogenic avian influenza virus. Similarly, the introduction of H5N1 virus into Vietnam, Thailand and Malaysia was estimated during March 2003, while that of the Indonesia it was during November 2002. The introduction of the H5N1 virus to Egypt was estimated to be September 2005 (Cattoli et al., 2011), which is approximately 5 months before the notification of the disease.
INDIAN SCENARIO
Subsequent to the outbreaks in Qinghai Lake (China), the H5N1 virus moved to South Asia over Himalayas, probably through migratory birds (Chen et al., 2005). In South Asia, the H5N1 virus was first detected in poultry in India during February 2006 from the State of Maharashtra (Pattnaik et al., 2006). Since then, India had reported a number of H5N1 outbreaks in poultry in several states including Manipur (July 2007), West Bengal and Tripura (January – May 2008), Assam, Sikkim and West Bengal (November 2008 – May 2009), West Bengal (January 2010), Tripura (February – March 2011), Assam and West Bengal (August – September 2011), Odisha, Meghalaya and Tripura (January – April 2012), Karnataka (September 2012) and recently in Bihar (March, 2013) and Chhattisgarh (August, 2013) (http: // www.oie.int / ). The H5N1 virus has also been isolated in other species including ducks [West Bengal and Assam (2008); Tripura (2008 and 2011), Odisha (2012)], Crows [Jharkhand (2011); Bihar (2012); Maharashtra (2012); Odisha (2012, 2014); Assam (2008)], Goose [Tripura (2008)] and Turkeys [Karnataka (2012)].
Molecular phylogeny of the viruses isolated from poultry in India identified two major clades; clade 2. 2 (isolated during 2006 to 2010) (Dubey et al., 2009; Tosh et al., 2011b) and clade 2. 3. 2. 1 (2011) (Nagarajan et al., 2012). Within clade 2. 2, the H5N1 viruses formed four distinct groups; 2006 and 2007 isolates formed two independent groups, 2008 – 2010 isolates formed one group except early 2008 virus isolated in Malda district, West Bengal (Tosh et al., 2011b). The phylogenetic analysis indicated two separate introductions of the H5N1 viruses into Malda district (West Bengal) during 2008. Nucleotide sequence analysis showed differential expression of PB1 – F2 protein in Indian H5N1 viruses. While most of the H5N1 viruses had complete expression of PB1 – F2, some of the viruses had premature termination (Nagarajan et al., 2010) or complete abolition either due to the presence of stop codon or mutation in the start codon (95ATG97→95ACG97) (Tosh et al., 2011b). The sequence analysis of the isolates revealed presence of multibasic amino acids at HA1-HA2 junction defining HPAI virus. Presence of amino acids Gln and Gly at positions 222 and 224 (H5 numbering), respectively in the HA indicate preferential binding to avian-specific receptors. The specific amino acid mutations observed in the NA (G l u 1 1 9 A l a and A s n 2 9 4 S e r) and M2 (S e r 3 1 A s n) genes revealed emergence of viruses resistant to oseltamivir (Chakrabarti et al., 2009; Tosh et al., 2011b) and amantadine (Tosh et al., 2011a) during 2008 and 2010, respectively in West Bengal. The PB2 amino acid alteration G l u 6 2 7 L y s, which is attributed to increased virulence of H5N1 viruses in mammals, is present in the majorities of the clade 2. 2 viruses. However, H5N1 viruses with wild-type PB2 were also co-circulating in India (Nagarajan et al., 2010; Tosh et al., 2011b). In 2011, H5N1 viruses belonging to a new fourth-order clade 2. 3. 2. 1 were isolated in chickens and ducks in Tripura (Nagarajan et al. 2012). Within clade 2. 3. 2. 1, viruses from Bangladesh and India isolated in 2011 grouped separately (100% confidence) from the viruses of Nepal isolated in 2010 indicating two separate introductions of the virus to South Asia. Within clade 2. 3. 2. 1, the viruses had undergone extensive genetic divergence leading to identification of 3 major subgroups (A, B and C), and the Indian isolates of 2011 belonged to subgroup A (Creanga et al., 2013).
CONCLUSION
The Asian strain of H5N1 virus (Gs / GD / 96 – like lineage) is continuing its spread, evolving into diverse genetic clades, causing outbreaks in poultry and occasionally transmitting to humans, and the pandemic threat of the virus is persisting. Intensive surveillance in wild birds as well as in poultry, pigs and peri-domestic birds is important to understand the spread and evolution of the virus. Rapid identification of newly emerging variants using genome-based detection methods along with phylogenetic analysis is useful in tracing the origin, transmission and emergence of reassortants as well as drug-resistant mutants. However, the wide spread of H5N1 virus and its ever changing nature make control program more difficult. Therefore, educational programs to enhance awareness on avian influenza, animal husbandry practices and biosecurity measures are important in control of the avian influenza.
REFERENCES
Abdel-Moneim AS, Afifi MA, El-Kady MF (2011). Genetic drift evolution under vaccination pressure among H5N1 Egyptian isolates. Virol. J. 8: 283.
http://dx.doi.org/10.1186/1743-422X-8-283
PMid:21651796 PMCid:PMC3146449
Alexander DJ (2007). An overview of the epidemiology of avian influenza. Vaccine 25: 5637 – 5644.
http://dx.doi.org/10.1016/j.vaccine.2006.10.051
PMid:17126960
Becker WB (1966). The isolation and classification of tern virus: influenza virus A/tern/South Africa/1961. J. Hyg. (Lond.) 64(3): 309 – 320.
http://dx.doi.org/10.1017/S0022172400040596
Boltz DA, Douangngeun B, Phommachanh P, Sinthasak S, Mondry R, Obert CA, Seiler P, Keating R, Suzuki Y, Hiramatsu H, Govorkova EA, Webster RG (2010). Emergence of H5N1 avian influenza viruses with reduced sensitivity to neuraminidase inhibitors and novel reassortants in Lao PDR. J. Gen. Virol. 91: 949 – 959.
http://dx.doi.org/10.1099/vir.0.017459-0
PMid:20016036 PMCid:PMC2888158
Bröjer C, Agren EO, Uhlhorn H, Bernodt K, Mörner T, Jansson DS, Mattsson R, Zohari S, Thorén P, Berg M, Gavier-Widén D (2009). Pathology of natural highly pathogenic avian influenza H5N1 infection in wild tufted ducks (Aythya fuligula). J. Vet. Diagn. Invest. 21(5):579 - 587.
http://dx.doi.org/10.1177/104063870902100501
PMid:19737752
Cattoli G, Fusaro A, Monne I, Coven F, Joannis T, El-Hamid HS, Hussein AA, Cornelius C, Amarin NM, Mancin M, Holmes EC, Capua I (2011). Evidence for differing evolutionary dynamics of A/H5N1 viruses among countries applying or not applying avian influenza vaccination in poultry. Vacc. 29(50): 9368 - 9375.
http://dx.doi.org/10.1016/j.vaccine.2011.09.127
PMid:22001877
CDC (1998) Update. Isolation of avian influenza A (H5N1) viruses from humans – Hong Kong, 1997 – 1998.Morb. Mortal. Wkly Rep. 46: 1245 – 1247.
PMid:9436713
Chakrabarti AK, Pawar SD, Cherian SS, Koratkar SS, Jadhav SM, Pal B, Raut S, Thite V, Kode SS, Keng SS, Payyapilly BJ, Mullick J, Mishra AC (2009). Characterization of the influenza A H5N1 viruses of the 2008-09 outbreaks in India reveals a third introduction and possible endemicity. PLoS One. 4(11): e7846.
http://dx.doi.org/10.1371/journal.pone.0007846
PMid:19924254 PMCid:PMC2775943
Chen H, Li Y, Li Z, Shi J, Shinya K, Deng G, Qi Q, Tian G, Fan S, Zhao H, Sun Y, Kawaoka y (2006a). Properties and dissemination of H5N1 viruses isolated during an influenza outbreak in migratory waterfowl in western China. J. Virol. 80:5976 – 5983.
http://dx.doi.org/10.1128/JVI.00110-06
PMid:16731936 PMCid:PMC1472608
Chen H, Smith GJ, Li KS, Wang J, Fan XH, Rayner JM, Vijaykrishna D, Zhang JX, Zhang LJ, Guo CT, Cheung CL, Xu KM, Duan L, Huang K, Qin K, Leung YH, Wu WL, Lu HR, Chen Y, Xia NS, Naipospos TS, Yuen KY, Hassan SS, Bahri S, Nguyen TD, Webster RG, Peiris JS, Guan Y (2006b). Establishment of multiple sublineages of H5N1 influenza virus in Asia: implications for pandemic control. Proc. Natl. Acad. Sci. USA 103: 2845 - 2850.
http://dx.doi.org/10.1073/pnas.0511120103
PMid:16473931 PMCid:PMC1413830
Chen H, Smith GJD, Zhang SY, Qin K, Wang J, Li KS, Webster RG, Peiris JSM, Guan Y (2005). Avian flu: H5N1 virus outbreak in migratory waterfowl. Nat. 436: 191 - 192.
http://dx.doi.org/10.1111/j.2005.0908-8857.03496.x
http://dx.doi.org/10.1111/j.0908-8857.2005.03496.x
Chen LM, Blixt O, Stevens J, Lipatov AS, Davis CT, Collins BE, Cox NJ, Paulson JC, Donis RO (2012). In vitro evolution of H5N1 avian influenza virus toward human-type receptor specificity. Virol. 422: 105 – 113.
http://dx.doi.org/10.1016/j.virol.2011.10.006
PMid:22056389
Cheung C-L, Rayner JM, Smith GJD, Wang P, Naipospos TSP, Zhang J, Yuen K-Y, Webster RG, Peiris JSM, Guan Y, Chen H (2006). Distribution of amantadine-resistant H5N1 avian influenza variants in Asia. J. Infect. Dis. 193: 1626 – 1629.
http://dx.doi.org/10.1086/504723
PMid:16703504
Cheung CL, Rayner JM, Wu W, Lau SY, Wang P, Tai H, Wen X, Song W, Mok B, Yuen KY, Peiris JSM, Guan Y, Chen H (2010). Naturally occurring antiviral drug resistance in avian H5N1 viruses. Health Research Symposium 2010, Poster presentations on Resp. Diseases, Abstract 143 (p85).
Cheung CL, Vijaykrishna D, Smith GJ, Fan XH, Zhang JX, Bahl J, Duan L, Huang K, Tai H, Wang J, Poon LL, Peiris JS, Chen H, Guan Y (2007). Establishment of influenza A virus (H6N1) in minor poultry species in southern China. J. Virol. 81(19): 10402 - 10412.
http://dx.doi.org/10.1128/JVI.01157-07
PMid:17652385 PMCid:PMC2045442
Creanga A, Nguyen DT, Gerloff N, Thi Do H, Balish A, Dang Nguyen H, Jang Y, Thi Dam V, Thor S, Jones J, Simpson N, Shu B, Emery S, Berman L, Nguyen HT, Bryant JE, Lindstrom S, Klimov A, Donis RO, Davis CT, Nguyen T (2013). Emergence of multiple clade 2.3.2.1 influenza A (H5N1) virus subgroups in Vietnam and detection of novel reassortants. Virology 444(1 - 2): 12 - 20.
Domenech J, Dauphin G, Rushton J, McGrane J, Lubroth J, Tripodi A, Gilbert J, Sims LD (2009). Experiences with vaccination in countries endemically infected with highly pathogenic avian influenza: the Food and Agriculture Organization perspective. Rev. Sci. Tech. 28:293 – 305.
PMid:19618633
Domingo E, Martínez-Salas E, Sobrino F, de la Torre JC, Portela A, Ortín J, López-Galindez C, Pérez-Bre-a P, Villanueva N, Nájera R, VandePol S, Steinhauer D, DePolo N, Holland JJ (1985). The quasispecies (extremely heterogeneous) nature of viral RNA genome populations: biological relevance-a review. Gene. 40(1):1 - 8.
http://dx.doi.org/10.1016/0378-1119(85)90017-4
Duan L, Bahl J, Smith GJ, Wang J, Vijaykrishna D, Zhang LJ, Zhang JX, Li KS, Fan XH, Cheung CL, Huang K, Poon LL, Shortridge KF, Webster RG, Peiris JS, Chen H, Guan Y (2008). The development and genetic diversity of H5N1 influenza virus in China, 1996 – 2006. Virology 380: 243 - 254.
http://dx.doi.org/10.1016/j.virol.2008.07.038
PMid:18774155 PMCid:PMC2651962
Dubey SC, Nagarajan S, Tosh C, Bhatia S, Lal Krishna. (2009). Avian influenza: A long known disease and its current threat. Indian J. Anim. Sci. 79: 113 - 140.
Earhart KC, Elsayed NM, Saad MD, Gubareva LV, Nayel A, Deyde VM, Abdelsattar A, Abdelghani AS, Boynton BR, Mansour MM, Essmat HM, Klimov A, Shuck-Lee D, Monteville MR, Tjaden JA (2009). Oseltamivir resistance mutation N294S in human influenza A (H5N1) virus in Egypt. J. Infect. Public Health 2: 74 – 80.
http://dx.doi.org/10.1016/j.jiph.2009.04.004
PMid:20701864
Ellis TM, Bousfield RB, Bissett LA, Dyrting KC, Luk GS, Tsim ST, Sturm-Ramirez K, Webster RG, Guan Y, Peiris JSM (2004). Investigation of outbreaks of highly pathogenic H5N1 avian influenza in waterfowl and wild birds in Hong Kong in late 2002. Avian Pathol. 33(5): 492 - 505.
http://dx.doi.org/10.1080/03079450400003601
PMid:15545029
Forrest HL, Webster RG (2010). Perspectives on influenza evolution and the role of research. Anim. Health Res. Rev. 11: 3 - 18.
http://dx.doi.org/10.1017/S1466252310000071
PMid:20591210
Fusaro A, Nelson MI, Joannis T, Bertolotti L, Monne I, Salviato A, Olaleye O, Shittu I, Sulaiman L, Lombin LH, Capua I, Holmes EC, Cattoli G (2010). Evolutionary Dynamics of Multiple Sublineages of H5N1 Influenza Viruses in Nigeria from 2006 to 2008. J. Virol. 84(7): 3239 – 3247.
http://dx.doi.org/10.1128/JVI.02385-09
PMid:20071565 PMCid:PMC2838112
Gerloff NA, Khan SU, Balish A, Shanta IS, Simpson N, Berman L, Haider N, Poh MK, Islam A, Gurley E, Hasnat MA, Dey T, Shu B, Emery S, Lindstrom S, Haque A, Klimov A, Villanueva J, Rahman M, Azziz-Baumgartner E, Ziaur Rahman M, Luby SP, Zeidner N, Donis RO, Sturm-Ramirez K, Davis CT (2014). Multiple reassortment events among highly pathogenic avian influenza A (H5N1) viruses detected in Bangladesh. Virol. 450 - 451: 297 - 307.
Govorkova EA, Baranovich T, Seiler P, Armstrong J, Burnham A, Guan Y, Peiris M, Webby RJ, Webster RG (2013). Antiviral resistance among highly pathogenic influenza A (H5N1) viruses isolated worldwide in 2002-2012 shows need for continued monitoring. Antiviral Res. 98(2): 297 - 304.
http://dx.doi.org/10.1016/j.antiviral.2013.02.013
PMid:23458714 PMCid:PMC3648604
Guan Y, Peiris JSM, Lipatov AS, Ellis TM, Dyrting KC, Krauss S, Zhang LJ, Webster RG, Shortridge KF (2002b). Emergence of multiple genotypes of H5N1 avian influenza viruses in Hong Kong SAR. Proc. Natl. Acad. Sci. USA 99(13): 8950 – 8955.
http://dx.doi.org/10.1073/pnas.132268999
PMid:12077307 PMCid:PMC124404
Guan Y, Peiris M, Kong KF, Dyrting KC, Ellis TM, Sit T, Zhang LJ, Shortridge KF (2002a). H5N1 Influenza Viruses Isolated from Geese in Southeastern China: Evidence for Genetic Reassortment and Interspecies Transmission to Ducks. Virol. 292: 16 - 23.
http://dx.doi.org/10.1006/viro.2001.1207
PMid:11878904
Guan Y, Shortridge KF, Krauss S, Webster RG (1999). Molecular characterization of H9N2 influenza viruses: were they the donors of the "internal" genes of H5N1 viruses in Hong Kong? Proc. Natl. Acad. Sci. USA 96:9363 – 9367.
http://dx.doi.org/10.1073/pnas.96.16.9363
PMid:10430948 PMCid:PMC17788
He G, Qiao J, Dong C, He C, Zhao L, Tian Y (2008). Amantadine-resistance among H5N1 avian influenza viruses isolated in Northern China. Antiviral Res. 77: 72 - 76.
http://dx.doi.org/10.1016/j.antiviral.2007.08.007
PMid:17897729
Hill AW, Guralnick RP, Wilson MJ, Habib F, Janies D (2009). Evolution of drug resistance in multiple distinct lineages of H5N1 avian influenza. Infect. Genet. Evol. 9(2): 169 - 178.
http://dx.doi.org/10.1016/j.meegid.2008.10.006
PMid:19022400
Höper D, Kalthoff D, Hoffmann B, Beer M (2012). Highly pathogenic avian influenza virus subtype H5N1 escaping neutralization: more than HA variation. J. Virol. 86(3): 1394 – 1404.
http://dx.doi.org/10.1128/JVI.00797-11
PMid:22090121 PMCid:PMC3264362
Hurt AC, Holien JK, Barr IG (2009). In vitro generation of neuraminidase inhibitor resistance in A (H5N1) influenza viruses. Antimicrob. Agents Chemother. 53: 4433 – 4440.
http://dx.doi.org/10.1128/AAC.00334-09
PMid:19651908 PMCid:PMC2764219
Hurt AC, Selleck P, Komadina N, Shaw R, Brown L, Barr IG (2007). Susceptibility of highly pathogenic A (H5N1) avian influenza viruses to the neuraminidase inhibitors and adamantanes. Antiviral Res. 73: 228 – 231.
http://dx.doi.org/10.1016/j.antiviral.2006.10.004
PMid:17112602
Ibrahim M, Eladl AF, Sultan HA, Arafa AS, Abdel Razik AG, Abd El Rahman S, El-Azm KI, Saif YM, Lee CW (2013). Antigenic analysis of H5N1 highly pathogenic avian influenza viruses circulating in Egypt (2006 – 2012). Vet. Microbiol. 167(3 - 4): 651 - 661.
Imai M, Kawaoka Y (2012). The role of receptor binding specificity in interspecies transmission of influenza viruses. Curr. Opin. Virol. 2(2): 160 - 167.
http://dx.doi.org/10.1016/j.coviro.2012.03.003
PMid:22445963
Kang HM, Batchuluun D, Kim MC, Choi JG, Erdene-Ochir TO, Paek MR, Sugir T, Sodnomdarjaa R, Kwon JH, Lee YJ (2011). Genetic analyses of H5N1 avian influenza virus in Mongolia, 2009 and its relationship with those of eastern Asia. Vet Microbiol. 147(1 - 2): 170 - 175.
Khan SU, Berman L, Haider N, Gerloff N, Rahman MZ, Shu B, Rahman M, Dey TK, Davis TC, Das BC, Balish A, Islam A, Teifke JP, Zeidner N, Lindstrom S, Klimov A, Donis RO, Luby SP, Shivaprasad HL, Mikolon AB (2013) Investigating a crow die-off in January-February 2011 during the introduction of a new clade of highly pathogenic avian influenza virus H5N1 into Bangladesh. Arch. Virol. 10.1007 / s00705 – 013 – 1842 - 0
Kilpatrick AM, Chmura AA, Gibbons DW, Fleischer RC, Marra PP, Daszak P (2006). Predicting the global spread of H5N1 avian influenza. Proc. Natl. Acad. Sci. USA 103(51): 19368 -19373.
http://dx.doi.org/10.1073/pnas.0609227103
PMid:17158217 PMCid:PMC1748232
Kou Z, Li Y, Yin Z, Guo S, Wang M, Gao X, Li P, Tang L, Jiang P, Luo Z, Xin Z, Ding C, He Y, Ren Z, Cui P, Zhao H, Zhang Z, Tang S, Yan B, Lei F, Li T (2009). The survey of H5N1 flu virus in wild birds in 14 Provinces of China from 2004 to 2007. PLoS One 4(9): e6926.
http://dx.doi.org/10.1371/journal.pone.0006926
PMid:19742325 PMCid:PMC2735031
Le QM, Kiso M, Someya K, Sakai YT, Nguyen TH, Nguyen KH, Pham ND, Ngyen HH, Yamada S, Muramoto Y, Horimoto T, Takada A, Goto H, Suzuki T, Suzuki Y, Kawaoka Y (2005). Avian flu: isolation of drug-resistant H5N1 virus. Nat. 437: 1108.
http://dx.doi.org/10.1038/4371108a
PMid:16228009
Li KS, Guan Y, Wang J, Smith GJ, Xu KM, Duan L, Rahardjo AP, Puthavathana P, Buranathai C, Nguyen TD, Estoepangestie AT, Chaisingh A, Auewarakul P, Long HT, Hanh NT, Webby RJ, Poon LL, Chen H, Shortridge KF, Yuen KY, Webster RG, Peiris JS (2004). Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nat. 430: 209 - 213.
http://dx.doi.org/10.1038/nature02746
PMid:15241415
Lipatov AS, Evseenko VA, Yen HL, Zaykovskaya AV, Durimanov AG, Zolotykh SI, Netesov SV, Drozdov IG, Onishchenko GG, Webster RG, Shestopalov AM (2007). Influenza (H5N1) Viruses in Poultry, Russian Federation, 2005 – 2006. Emerg. Infect. Dis. 13(4): 539 - 546.
http://dx.doi.org/10.3201/eid1304.061266
PMid:17553267 PMCid:PMC2725974
Matrosovich M, Zhou, N, Kawaoka, Y, Webster R (1999). The surface glycoproteins of H5 influenza viruses isolated from humans, chickens, and wild aquatic birds have distinguishable properties. J. Virol. 73: 1146 – 1155.
PMid:9882316 PMCid:PMC103935
Monne I, Fusaro A, Al-Blowi MH, Ismail MM, Khan OA, Dauphin G, Tripodi A, Salviato A, Marangon S, Capua I, Cattoli G (2008). Co-circulation of two sub-lineages of HPAI H5N1 virus in the Kingdom of Saudi Arabia with unique molecular signatures suggesting separate introductions into the commercial poultry and falconry sectors. J. Gen. Virol. 89: 2691 – 2697.
http://dx.doi.org/10.1099/vir.0.2008/004259-0
PMid:18931064 PMCid:PMC2886959
Monne I, Yamage M, Dauphin G, Claes F, Ahmed G, Giasuddin M, Salviato A, Ormelli S, Bonfante F, Schivo A, Cattoli G (2013). Reassortant avian influenza A (H5N1) viruses with H9N2-PB1 gene in poultry, Bangladesh. Emerg. Infect. Dis. 19 (10): 1630 - 1634.
http://dx.doi.org/10.3201/eid1910.130534
PMid:24047513 PMCid:PMC3811991
Nagarajan S, Tosh C, Murugkar HV, Venkatesh G, Katare M, Jain R, Behera P, Khandia R, Tripathi S, Kulkarni DD, Dubey SC (2010). Isolation and molecular characterization of a H5N1 virus isolated from a Jungle crow (Corvus macrohynchos) in India. Virus Genes. 41(1): 30 - 36.
http://dx.doi.org/10.1007/s11262-010-0477-4
PMid:20396941
Nagarajan S, Tosh C, Smith DK, Peiris JS, Murugkar HV, Sridevi R, Kumar M, Katare M, Jain R, Syed Z, Behera P, Cheung CL, Khandia R, Tripathi S, Guan Y, Dubey SC (2012). Avian influenza (H5N1) virus of clade 2.3.2 in domestic poultry in India. PLoS One. 7(2): e31844.
http://dx.doi.org/10.1371/journal.pone.0031844
PMid:22363750 PMCid:PMC3282738
Pattnaik B, Pateriya AK, Khandia R, Tosh C, Nagarajan S, Gounalan S, Murugkar HV, Shankar BP, Shrivastava N, Behera P, Bhagat S, Peiris JSM, Pradhan HK (2006). Phylogenetic analysis revealed genetic similarity of the H5N1 avian influenza viruses isolated from HPAI outbreaks in chickens in Maharashtra, India with those isolated from swan in Italy and Iran in 2006. Curr. Sci. 91: 77 – 81.
Peiris JS, Yu WC, Leung CW, Cheung CY, Ng WF, Nicholls JM, Ng TK, Chan KH, Lai ST, Lim WL, Yuen KY, Guan Y (2004). Re-emergence of fatal human influenza A subtype H5N1 disease. Lancet. 363 (9409): 617 - 619.
http://dx.doi.org/10.1016/S0140-6736(04)15595-5
Pfeiffer DU, Otte MJ, Roland-Holst D, Inui K, Nguyen T, Zilberman D (2011). Implications of global and regional patterns of highly pathogenic avian influenza virus H5N1 clades for risk management. Vet. J. 190(3): 309 - 316.
http://dx.doi.org/10.1016/j.tvjl.2010.12.022
PMid:21288747
Reid SM, Shell WM, Barboi G, Onita I, Turcitu M, Cioranu R, Marinova-Petkova A, Goujgoulova G, Webby RJ, Webster RG, Russell C, Slomka MJ, Hanna A, Banks J, Alton B, Barrass L, Irvine RM, Brown IH (2011). First Reported Incursion of Highly Pathogenic Notifiable Avian Influenza A H5N1 Viruses from Clade 2.3.2 into European Poultry. Transbound. Emerg. Dis. 58: 76 – 78.
http://dx.doi.org/10.1111/j.1865-1682.2010.01175.x
PMid:21054819 PMCid:PMC3582342
Salzberg SL, Kingsford C, Cattoli G, Spiro DJ, Janies DA, Aly MM (2007). Genome analysis linking recent European and African influenza (H5N1) viruses. Emerg. Infect. Dis. 13: 713 – 718.
http://dx.doi.org/10.3201/eid1305.070013
PMid:17553249 PMCid:PMC2432181
Savill NJ, St Rose SG, Keeling MJ, Woolhouse ME (2006). Silent spread of H5N1 in vaccinated poultry. Nature 442: 757.
http://dx.doi.org/10.1038/442757a
PMid:16915278
Sharshov K, Silko N, Sousloparov I, Zaykovskaya A, Shestopalov A, Drozdov I (2010). Avian influenza (H5N1) outbreak among wild birds, Russia, 2009. Emerg. Infect. Dis. 16: 349 – 351.
http://dx.doi.org/10.3201/eid1602.090974
PMid:20113582 PMCid:PMC2958015
Sitaras I, Kalthoff D, Beer M, Peeters B, de Jong MC (2014). Immune escape mutants of Highly Pathogenic Avian Influenza H5N1 selected using polyclonal sera: identification of key amino acids in the HA protein. PLoS One. 9(2): e84628.
http://dx.doi.org/10.1371/journal.pone.0084628
PMid:24586231 PMCid:PMC3934824
Smith GJD, Fan XH, Wang J, Li KS, Qin K, Zhang JX, Vijaykrishna D, Cheung CL, Huang K, Rayner JM, Peiris JS, Chen H, Webster RG, Guan Y (2006). Emergence and predominance of an H5N1 influenza variant in China. Proc. Natl. Acad. Sci. USA 103: 16936 – 16941.
http://dx.doi.org/10.1073/pnas.0608157103
PMid:17075062 PMCid:PMC1636557
Smith GJ, Vijaykrishna D, Ellis TM, Dyrting KC, Leung YH, Bahl J, Wong CW, Kai H, Chow MK, Duan L, Chan AS, Zhang LJ, Chen H, Luk GS, Peiris JS, Guan Y (2009). Characterization of avian influenza viruses A (H5N1) from wild birds, Hong Kong, 2004 -2008. Emerg. Infect. Dis. 15(3): 402 - 407.
http://dx.doi.org/10.3201/eid1503.081190
PMid:19239752 PMCid:PMC2666293
Starick E, Beer M, Hoffmann B, Staubach C, Werner O, Globig A, Strebelow G, Grund C, Durban M, Conraths FJ, Mettenleiter T, Harder T (2008). Phylogenetic analyses of highly pathogenic avian influenza virus isolates from Germany in 2006 and 2007 suggest at least three separate introductions of H5N1 virus. Vet. Microbiol. 128(3-4): 243 - 252.
http://dx.doi.org/10.1016/j.vetmic.2007.10.012
PMid:18031958
Suarez DL (2000). Avian influenza: our current understanding. Anim. Health. Res. Rev. 11:19 – 33.
http://dx.doi.org/10.1017/S1466252310000095
PMid:20591211
Suarez DL, Perdue ML, Cox N, Rowe T, Bender C, Huang J, Swayne DE (1998). Comparisons of Highly Virulent H5N1 Influenza A Viruses Isolated from Humans and Chickens from Hong Kong. J. Virol. 72(8): 6678 - 6688.
PMid:9658115 PMCid:PMC109865
Subbarao K, Klimov A, Katz J, Regnery H, Lim W, Hall H, Perdue M, Swayne D, Bender C, Huang J, Hemphill M, Rowe T, Shaw M, Xu X, Fukuda K, Cox N (1998). Characterization of an avian influenza A (H5N1) virus isolated from a child with a fatal respiratory illness. Science 279: 393 – 396.
http://dx.doi.org/10.1126/science.279.5349.393
PMid:9430591
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013). MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 30: 2725 - 2729.
http://dx.doi.org/10.1093/molbev/mst197
PMid:24132122
Teifke JP, Klopfleisch R, Globig A, Starick E, Hoffmann B, Wolf PU, Beer M, Mettenleiter TC, Harder TC (2007). Pathology of natural infections by H5N1 highly pathogenic avian influenza virus in mute (Cygnus olor) and whooper (Cygnus Cygnus) swans. Vet. Pathol. 44(2): 137 - 143.
http://dx.doi.org/10.1354/vp.44-2-137
PMid:17317790
Tosh C, Murugkar HV, Nagarajan S, Tripathi S, Katare M, Jain R, Khandia R, Syed Z, Behera P, Patil S, Kulkarni DD, Dubey SC (2011a). Emergence of amantadine-resistant avian influenza H5N1 virus in India. Virus Genes. 42: 10 - 15.
http://dx.doi.org/10.1007/s11262-010-0534-z
PMid:20953687
Tosh C, Nagarajan S, Murugkar HV, Jain R, Behera P, Katare M, Kulkarni DD, Dubey SC (2011b) Phylogenetic evidence of multiple introduction of H5N1 virus in Malda district of West Bengal, India in 2008. Vet. Microbiol. 148(2 - 4): 132 - 139.
Tran DL, Kim K, Choi JY, Paik HD, Choi SW, Ma JY, Kim SS, Ahn SJ, Kim YB (2011). Genome sequence analysis of H5N1 influenza a virus isolated from a Vietnamese in 2007. J. Microbiol. 49(2): 274 - 279.
http://dx.doi.org/10.1007/s12275-011-0311-2
PMid:21538249
Vijaykrishna D, Bahl J, Riley S, Duan L, Zhang JX, Chen H, Peiris JSM, Smith GJD, Guan Y (2008). Evolutionary Dynamics and Emergence of Panzootic H5N1 Influenza Viruses. PLoS Pathol. 4(9): e1000161.
http://dx.doi.org/10.1371/journal.ppat.1000161
PMid:18818732 PMCid:PMC2533123
Wang ZG, Jiang WM, Liu S, Hou GY, Li JP, Wang ZY, Chen JM (2012). Increased substitution rate in H5N1 avian influenza viruses during mass vaccination of poultry. Chin. Sci. Bull. 57: 2419 - 2424.
http://dx.doi.org/10.1007/s11434-012-5215-y
WHO (2014) Antigenic and genetic characteristics of zoonotic influenza viruses and development of candidate vaccine viruses for pandemic preparedness.(http://www.who.int/influenza/resources/documents/characteristics_virus_vaccines/en/, accessed on 23 April 2014)
WHO/FAO/OIE H5N1 Evolution Working Group (2008). Towards a unified nomenclature system for the highly pathogenic avian influenza H5N1 viruses. Emerg. Infect. Dis. 14, In: http://www.cdc.gov/EID/content/14/7/e1.html., WHO/OIE/FAO H5N1 Evolution Working Group (2012). Continued evolution of highly pathogenic avian influenza A (H5N1): Updated nomenclature. Influenza Other Respi. Viruses 6(1): 1 – 5.
WHO/OIE/FAO H5N1 Evolution Working Group (2014). Revised and updated nomenclature for highly pathogenic avian influenza A (H5N1) viruses. Influenza Other Respi. Viruses DOI: 10.1111/irv.12230.
http://dx.doi.org/10.1111/irv.12230
Xiao X, Zhang WD, Duan BF, Zhao HY, Liu QL, Hu TS, Qiu W, Feng ZL, Zheng Y, Fan QS, Zhang YG, Zhang FQ (2013). Genetic evolution analysis of matrix protein 2 gene of avian influenza H5N1 viruses from boundary of Yunnan province. Zhonghua Yu Fang Yi Xue Za Zhi. 47(6): 514 - 517.
PMid:24113099
Xu X, Subbarao K, Cox NJ, Guo Y (1999). Genetic characterization of the pathogenic influenza A/Goose/Guangdong/1/96 (H5N1) virus: similarity of its hemagglutinin gene to those of H5N1 viruses from the 1997 outbreaks in Hong Kong. Virol. 261: 15 – 19.
http://dx.doi.org/10.1006/viro.1999.9820

