
Advances in Animal and Veterinary Sciences


Review Article
Advances in Animal and Veterinary Sciences 2 (2S): 46 – 54Special Issue 2 (2014) (Advances in Diagnosis and Control of Infectious Diseases of Animals)
Equine Herpesviruses: a Brief Review
Sanjay Kapoor1, Himanshu Sharma1*, Mahavir Singh1, Pankaj Kumar1, Koushlesh Ranjan2, Anuradha Kumari3, Richa Khirbat4
- Department of Veterinary Microbiology, LLR University of Veterinary and Animal Sciences, Hisar, Haryana, India, 125004
- Department of Veterinary Physiology and Biochemistry, Sardar Vallabhbhai Patel University of Agriculture and Technology, Meerut, Uttar Pradesh, India, 250110
- Department of Livestock Production Management, LLR University of Veterinary and Animal Sciences, Hisar, Haryana, India, 125004
- Haryana Veterinary Vaccine Institute, Hisar, Haryana, India, 125001.
*Corresponding author:himshu2002@yahoo.co.in
ARTICLE CITATION:
Kapoor S, Sharma H, Singh M, Kumar P, Ranjan K, Kumari A, Khirbat R (2014). Equine herpesviruses: a brief review. Adv. Anim. Vet. Sci. 2 (2S): 46 – 54.
Received: 2014–05–26, Revised: 2014–06–25, Accepted: 2014–06–27
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/journal.aavs/2014/2.2s.46.54
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Nine equine herpesviruses (EHV) have been known to infect equines. Equine herpesvirus type–1 (EHV1) is an important and ubiquitous viral pathogen of the horse and produces syndromes of respiratory disease, epidemic abortion and sporadic encephalomyelitis. This review article is focused on some aspects of EHV1 biology, its life cycle and pathogenicity in the natural host. Vaccination is one of the best options to fight EHV1 infection. Various strategies of vaccination that have been investigated and developed over the past decades will be presented in this review. Diagnosis is important to start specific treatment. The latest diagnostic techniques which were developed recently together with conventional will also be discussed.
INTRODUCTION
Equine herpesviruses (EHVs) belong to the family Herpesviridae; a term derived from a latin word which means ‘to creep’. The family is subdivided into three subfamilies (a B and y) based on tissue tropism, pathogenicity and behavior in tissue culture (McGeoch et. al.,2000; Davison et. al.,2002). Nine species of EHV have been reported (Table 1). The species EHV1, EHV3, EHV4, EHV8 and EHV9 have been classified under the genus Varicellovirus, subfamily Alphaherpesvirinae, family Herpesviridae of the order Herpesvirales. The species EHV2 and EHV5 have been put under a new genus Percavirus, subfamily Gamaherpesvirinae, family Herpesviridae of the order Herpesvirales. The EHV6 and EHV7 have been tentatively placed as species in the subfamily Alphaherpesvirinae and Gammaherpesvirinae, respectively (Davison at el., 2009). Only five of the nine herpes viruses (viz. EHV1, 2, 3, 4 and 5) have the ability to produce disease in horses. EHV3 is responsible for coital exanthema while EHV2 and 5 are not associated with a specific disease but may remain associated with upper respiratory tract diseases, inappetance, lymphadenopathy, immunosupression, keratoconjunctivitis, general malaise and poor performance (Thein et al. 1978; Plummer et al.1973; Belak et al. 1980). Both EHV1 and 4 are economically important viruses affecting the respiratory tracts of horses globally (Davison et. al., 2009; Roizman, 1996; Allen and Bryans, 1986; Brosnahan and Osterrieder, 2009). However, only EHV1 causes abortion and neurological disorders (Patel and Heldens, 2005; Lunn et. al., 2009). It was only in 1981 that the EHV1 and EHV4 were considered antigenically and genetically two distinct viruses, and not the same virus, based on DNA fingerprint analysis (Sabine et. al., 1981; Studdert et. al., 1981). This review will mainly focus EHV1 together with EHV4.

VIRUS STRUCTURE
The equine herpes virions with a diameter of ~150 nm have icosahedral nucleocapsid surrounded by a layer of globular tegument which in turn is enclosed in an envelope. Several glycoprotein peplomers are present on the viral envelope. The genome of equine herpes virions is made up of single linear dsDNA molecule with 57% G+C ratio. The genome size slightly varies between EHV1 (150 kbp) and EHV4 (145 kbp) (Darlington and Randall, 1963; Roizman et. al., 1992). Another feature of the genomes EHV1 and EHV4 is that they are composed of a unique long (UL) region attached to a unique short (US) region. A set of indirect repeat sequences called internal and terminal sequences (IRS and TRS) flank the US (Figure 1) (Telford et. al., 1992, 1994). The UL, US and IRS regions have 63, 9, and 4 genes, respectively, out of the total 76 genes reported in EHV1 and EHV4. Well–controlled expression of the genes divided into three distinct phases viz. Immediate Early (IE), Early (E), and Late (L) is observed (Caughman et. al., 1985, Albrecht et. al., 2005). Whole genome sequencing (Teleford et. al., 1992, 1998) of EHV1 and EHV4 indicates that the genome actually has 80 and 79 open reading frames (ORFs), respectively, due to duplication of some genes. Various proteins of EHV1 and EHV4 have been found to be identical at amino acid sequence homology in the range of 55% to 96% (Teleford et. al., 1998).

Figure 1: EHV1 genome: a unique long (UL) region attached to a unique short (US) region. A set of indirect repeat sequences called internal and terminal sequences (IRS and TRS) flank the US
PATHOGENESIS
Respiratory Form
The binding of the EHV1 and EHV4 to heparan sulfate present on the surface of cell and subsequent interaction with a putative glycoprotein D receptor(s) leads to initiation of infection, (Azab and Osterrieder, 2012). Subsequent to attachment, entry of virus takes place either by receptor mediated endocytosis or direct fusion of the virus envelope with the plasma membrane. It has been observed that multiple endocytic pathways are utilized by EHV1 in different cell types to establish productive infection (El–Hasebe et. al., 2009). Equine brain microvascular endothelial cells are infected via caveolar endocytosis, while equine dermal cells involve energy– and pH–dependent endocytosis. After removal of the viral envelope following fusion, the naked virus particles start their replication in the nucleus after travelling through the cytoplasm. The integrity of the microtubule network and the minus–end microtubule motor protein, dynein plays an important role in the intracellular trafficking resulting in EHV1 infection (Frampton et. al., 2010).
After inhalation, the spread of EHV1 and EHV4 to regional lymph nodes occurs after their multiplication in the epithelium of respiratory tract (nasal cavities, pharynx, trachea, bronchi and bronchiole). Intranuclear inclusion bodies along with necrosis have been reported in respiratory epithelia and lymphoid germinal centres. A murine respiratory model has been used to study the pathogenesis of EHV1–induced respiratory disease in the horse (Awan et. al., 1991; Walker et. al., 1999). The role of proinflammatory beta chemokines in the pathogenesis of EHV1 have also been demonstrated (Smith et. al., 2000). Recently, pulmonary fibrosis in horses could experimentally be induced with the EHV5 (Williams mail et. al., 2013).
Abortion and Neonatal Disease
Abortion can be the result of exogenous or endogenous infection, i.e. reactivation of latent virus (Allen et. al., 1998). EHV1 virus after infecting respiratory tract spreads quickly throughout the body by invading leucocytes, lamina propria and endothelial cells of blood and lymphatic vessels due to a cell–to–cell spread of infectious virus from the respiratory epithelium.
Cell–associated viraemia is the result of movement of infected mononuclear cells from the regional lymph nodes into the circulation. The virus then cross the placenta, infect fetus and ultimately leads to abortion (Bryans et. al., 1969). EHV1 replicates in endothelial cells and induces thrombosis and ischaemia in the microcotyledons of the placenta. This was found to be the primary cause of abortion (Edington et. al., 1991; Smith et. al., 1992). However, not all EHV1 isolates have equal potential to cause abortion (Mumford et. al., 1994). Abortions induced by EHV1 don’t affect the mare’s subsequent reproductive efficiencies (Crabb et. al., 1996). Foetus may be born alive if infected in later stages of pregnancies, but soon after birth it dies because of respiratory distress, pneumonia and other respiratory complications (Crabb et. al., 1996).
Neurological Disease
EHV1 is non–neurotropic, even after intracerebral inoculation, in contrast to several other alphaherpesviruses (Jackson et. al., 1977; Prickett, 1969). Induction of myeloencephalitis reflects endotheliotropism but not neurotropism (Edington et. al., 1986; Jackson et. al., 1977; Patel et. al., 1982; Thein et. al., 1993; Whitwell and Blunden, 1992; Wilson, 1997). The minor blood vessels in the brain or spinal cord exhibit thrombosis and vasculitis due to infection of endothelial cells. However, multiplication of EHV1 strains in neurons and glial cells is observed after intracerebral inoculation of baby mice (Nowotny, 1987) resulting in neuronal latency in mice (Marshall and Field, 1997). Inbred mouse strains have proved to be very valuable models for studying the pathogenesis of EHV1 induced myeloencephalopathy in horses (Mori et. al., 2012). Recently, in suckling hamster model, it was demonstrated that EHV9 invade the brain via the trigeminal nerve besides the abducens, oculomotor, and facial nerves, which suggests the neuronal spread of neuropathogenic viruses to the brain via the eyes (Habashi et. al., 2013). Macroscopic lesions in the CNS include focal haemorrhage in parenchymal tissues of the brain and spinal cord which are not found frequently. Congestion, vasculitis, thrombosis and secondary ischemic degeneration are some of the significant histological lesions found in the CNS (Wilson, 1997). The requirement of horses as experimental animals to investigate EHV1 neuropathogenicity can be reduced with the use of equine respiratory explants for in vitro studies (Annelies et. al., 2010) with promising and reproducible results.
The neuropathogenicity of EHV1 strains is significantly associated with single nucleotide polymorphism (A2254 → G2254) in the open reading frame (ORF) 30, of the viral DNA polymerase, resulting in an amino acid variation from aspargine to aspartic acid (N/D752) (Figure 2) (Nugent et. al.,2006; Van de Walle et. al.,2009). How EHV1 neuropathogenic phenotype is influenced by the DNA polymerase genotype requires further studies (Pronost et. al., 2010). The site directed mutagenesis has been applied to further investigate and confirm relationship between single nucleotide polymorphism in EHV1 polymerase and neuropathogenicity (Goodman et. al., 2007).
Figure 2: Single nucleotide polymorphism in EHV1 DNA polymerase gene: nucleotide substitution (A2254 to G2254) in polymerase gene leads to conversion of abortive strain to neuropathogenic strain
LATENCY
EHV1 after infection produce latent infection in over 50% cases in neuronal and lymphoid tissues (Pusterla et. al., 2012) and persists for the life time in the host (Foote et. al., 2006). Latent infection can be reactivated by stress, corticosteroids or drugs (Browning et. al., 1988; Nageshi et. al., 1992; Barrandeguy et. al., 2008). Prototype of alphaherpesviruses i.e. herpes simplex virus establish latent infection in neuronal tissues (Stevens, 1989; Stevens, 1994). In contrast to it, EHV1 settles its latent infection in neuronal as well as lymphoid tissues (Edington et. al., 1985; Edington, 1994; Welch et. al., 1992). During latency entire viral genome persists in the infected cell but only its limited part transcribe in to latency associated transcripts (LATs), i.e. the only detectable RNA during latency. In EHV1 two LATs have been identified, derived from immediate early genes (gene 63 and 64) (Baxi et. al., 1995; Chester et. al., 1997; Borchers et. al., 1999). Targeting these LATs, latency can be detected by in situ hybridization, northern blotting, reverse transcriptase polymerase chain reaction (RT–PCR) or by real–time PCR (Stroop et. al., 1984; Stevens et. al., 1987; Pusterla et. al., 2009). Latency also has been identified as the absence of detectable expression of viral gene (ORF33) that encodes the late structural protein B (gB) (Allen et. al., 2008).
CLINICAL SIGNS AND SYMPTOMS
Respiratory Disease
The respiratory disease of equines can be caused by EHV1 as well as EHV4 (Allen et. al., 1986). Both viruses generally lead to mild or subclinical infection. The incubation period ranges from 2 to 10 days. The respiratory signs for EHV1 and EHV4 include high fever of 102 –107º F for 1–7 days, inappetence, depression, nasal discharge and coughing. The clinical signs are often mild or even absent after subsequent re–infection.
Abortion and Neonatal Disease
EHV1 is a leading cause of abortion in horses worldwide. Incubation period is highly variable between nine days to four months (Allen et. al., 1998). Abortion usually occurs after 2–12 weeks of infection in late gestation (between 7 and 11 months of gestation). The infection doesn’t cause damage to mare's reproductive tract, and she can conceive later. Mares if infected late in gestation period give birth to live foal but it can die within a few days after birth (Perkins et. al., 1999).
Encephalomyelitis
In recent years increased incidence of Equine herpesvirus myeloencephalopathy (EHM), has been observed by infection with some virulent strains of EHV1 leading to inflammation of the blood vessels that supply the brain and spinal cord (Henninger et. al.,2007). The neurological signs of Equine herpesvirus myeloencephalopathy (EHM) includes multifocal myeloencephalopathy, ischemic neuronal injury, hemorrhage andthrombosis (Wilson, 1997). The incubation period of encephalomyelitis is of 6–10 days. The onset of the neurologic signs is sudden and can vary from mild in–coordination and posterior paresis to severe posterior paralysis with recumbency, altered gaits, ataxia, staggering, inability to rise from the sitting position, loss of anal and bladder tone, and urinary incontinence. On rare occasions the paralysis may advance to quadriplegia and finally death of the animal (Studdert et. al., 2003, Goehring et. al., 2005).
TRANSMISSION
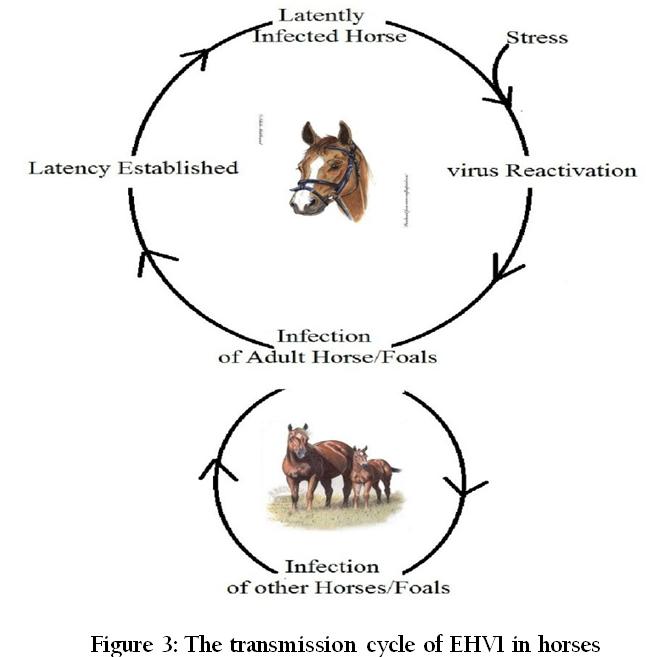
EHV1 is commonly referred to as the equine abortion virus. EHV4 is most common among foals and yearlings and, is also referred as equine rhinopneumonitis virus. The principal reservoir of infection for EHV is latently infected horses. It usually survives for less than 7 days in the environment, but has been detected for up to 35 days and is easily inactivated by heat and disinfectants. Equine herpesvirus spreads directly from horse–to–horse and indirectly by inanimate objects by coming in contact with nasal secretions and aborted fetuses or fetal fluids and aerosol from coughing horses. It is mainly transmitted by inhalation of the infected droplets or by the ingestion of material contaminated with nasal secretions, aborted fetuses, and placental membranes (Hebia et. al., 2007). Latent infection and its reactivation play an important role in the epidemiology of EHV1 abortion and neurologic disease (Figure 3).
PREVENTION
Vaccination is an effective tool to prevent EHV1 infection together with good managing practices (Kapoor et. al., 2010). Successful vaccination against EHV1 require both humoral as well as cell mediated immune response in a balanced proportion. Various commercialized vaccines can be categorized in to live attenuated, inactivated, subunit, recombinant and DNA vaccines. However, inactivated EHV1 vaccine is main vaccine which provides the variable level of protection against the disease.
Schedule of Vaccination
Two vaccinations one month apart are required to induce immunity, followed by boosters at every six months. Vaccination is recommended for foals over 3–5 months of age. It is best not to vaccinate horses when they are sick. To avoid EHV1 induced abortion, it is recommended to vaccinate pregnant mares at the 5th, 7th and 9th months of pregnancy.
INERT VACCINES
Inactivated whole EHV1 Vaccines and Sub–Unit Vaccines
EHV1 vaccines were first introduced in 1960. However, majority of the commercial vaccines are either inactivated whole virus vaccine or subunit vaccine. The major advantages of these vaccines are high degree of safety, as the virus replication and subsequent spread between hosts have been blocked. Viruses are usually inactivated by formaldehyde or β– propiolactone. Nowadays, inactivated whole EHV1 vaccines are widely used. In foals, vaccination significantly reduced symptoms of disease and the time–course of both cell–associated viremia and shedding of virus after experimental infection. In a recent study, ponies vaccinated twice with a conventional whole EHV1 inactivated vaccine adjuvanted with carbomer developed a strong humoral response (both CF and VN antibodies) after both immunizations (Minke et. al., 2006).
Despite an extensive use of inactivated virus vaccines in many countries over decades, abortion storms are still being reported (van Maanen et. al., 2000). Presently, whole virus inactivated vaccines have been replaced by subunit or split vaccines. The split or subunit vaccine is produced by disintegration of purified virion particles usually by chemical treatment. They generally induce similar adverse reactions because they contain the same constituents as inactivated whole virus vaccines. Due to the large number of proteins expressed by EHV1, the selection of relevant proteins for subunit EHV1 vaccines tends to be difficult. Glycoproteins of EHV1 are essential for cell receptor recognition and the entry into target cells. Glycoproteins B and C contain antibody epitopes strongly recognised by the immune system of horses infected or vaccinated with EHV1, and are therefore candidates for inclusion in sub–unit vaccines against EHV1 (Tewari et. al.,1995). Recombinant baculoviruses having gD and gH genes induce serum antibodies to EHV1 (Fuentealba et. al., 2014). Similar results have been obtained with baculovirus recombinant gC and gB (Kukreja et. al., 1998; Tewari et. al., 1995).
Immunostimulating Complex–Based Vaccines
Immunizations with proteins generally require the presence of strong adjuvants to stimulate an immune response. The Quillaia saponin (Quil A) are generally used as adjuvant and membrane protein/antigens are usually integrated in immuno–stimulating complexes (ISCOM™) or mixed with Iscomatrix™ to improve their antigenicity. ISCOMs and Iscomatrix™ are not fusogenic but can associate with intracellular lipid membranes in the cytoplasm and vesicular compartments (Bungener et. al., 2002).
An experimental sub–unit vaccine containing glycoproteins gp2, gp10, gB, gC, gD and gM, of EHV1 and adjuvant Iscomatrix™ showed antibody response with virus neutralization (VN) activity in hamsters (Cook et. al., 1990). A similar vaccine composed of inactivated Sf9 insect cells infected by a recombinant baculovirus coding for EHV1 gD and mixed with the adjuvant Iscomatrix™ was administered to ponies, pregnant mares and foals via the intramuscular route (Foote et. al., 2005). Vaccinated ponies developed a virus–specific antibody (having IgGa, IgGb, IgG (T) and IgA antibodies) response with neutralizing activity (Weerasinghe et. al., 2006) which was similar to the response induced by inactivated whole virus EHV1 vaccine.
Mares and foals vaccinated twice with a sub–unit vaccine containing both gB and gD mixed with the Iscomatrix™ adjuvant have shown reduced virus shedding. However, cell–associated viremia was not affected by vaccination (Foote et. al., 2006).
DNA Vaccines
DNA vaccine consists of plasmids expressing gene of whole antigenic proteins or simply epitopes of these proteins. These antigens are directly expressed in vivo in the target host cell and ultimately result in stimulation of both humoral as well as cellular immune responses. DNA vaccines can also be adjuvanted by the insertion of genes coding for co–stimulatory molecules (e.g. cytokines), theoretically allow a more controlled modulation of the immunity than the one induced by conventional adjuvant vaccines. DNA vaccines can be lyophilized; hence they have good stability and long–term storage. A DNA vaccine encoding gD has been prepared which elicit both cell–mediated as well as humoral immune responses (Ruitenberg et. al., 1999). An increased level of gD–specific antibody was observed in horses after intramuscular administration of DNA vaccine encoding gD (Ruitenberg et. al., 2000). The DNA vaccine containing coding genes of gB, gC and gD glycoproteins of EHV1 has been tested. DNA constructs were administrated alone, in combination with adjuvants (i.e. aluminum phosphate or carbopol), or complexed with the cationic lipid DMRIE–DOPE in the presence of a DNA plasmid coding for granulocyte–macrophage colony stimulating factor (GMCSF). The amount of VN antibody was increased in ponies vaccinated with the DNA/DMRIE–DOPE vaccine, and the duration of nasopharyngeal virus excretion after experimental infection with EHV1 was reduced following immunization with a DNA vaccine adjuvanted with aluminium phosphate (Minke et. al., 2006). It is clear that DNA vaccination efficiently stimulates an EHV1 specific antibody response in the horse but so far no cellular immune responses have been studied. Therefore, the protection afforded by this type of vaccine needs further investigation.
Non Infectious EHV1 L–Particles
The generation of non–infectious virion–related particles in cells shortly after infection with EHV1 has been carried out (McLauchlan and Rixon, 1992). They are comparatively less known type of virus–like particles formed by tegument proteins surrounded by an envelope and lack the internal capsid protein and viral nucleic acid. These L–particles do not contain viral DNA which makes them noninfectious. However, their capability to transport viral envelope proteins to target cells remains unaffected, and thus has no effect on their biological competence (Pardoe and Dargan, 2002). L–particles possess an intact envelope membrane with all immunogenic glycoproteins so they are expected to stimulate an immune response towards the viral glycoproteins similar to that during the natural viral infection. As a result, they have been identified as potential immunising agents. They are named light–particles (L–particles) because of their ability to be separated into a low density band on Ficoll gradients (Dargan and Subak–Sharpe, 1997). L–particles could support initiation of viral replication at the early stage of infection and/or may serve as decoys for the immune response. However, the exact role of L–particles in infection has not been established. In specific–pathogen–free (SPF) foals, immunization with EHV1 L–particles induced a CF antibody response. Clinical signs of disease, virus shedding and cell–associated viremia were milder in immunized ponies when compared with unvaccinated animals.
A new type of virus–related particle, called pre–viral DNA replication enveloped particles (PREPs), has been developed (Dargan et. al., 1995). The relative protein composition and structure of these PREPs is different from L–particles, though these two have similar morphology.
LIVE EHV1 VACCINES
Live vaccine contains live microorganisms such as live attenuated or a genetically modified organisms, or heterologous organism used as vector for the gene coding pathogen’s antigen.
Live Attenuated Vaccines
The immunization with live attenuated virus is very similar to natural infection. The intranasal or intramuscular inoculation of a live attenuated/modified EHV1 is an attractive approach for the vaccination against EHV1. However, the risk of reversion to virulence remains a concern for this strategy of vaccination. Two principal types of EHV1 mutants have been derived and used as live attenuated EHV1 vaccines, namely thymidine kinase negative (TK–) and temperature sensitive (Ts) mutants. Their virulence and ability to establish latency is reduced in the horse while immunogenicity and stability is retained. Immunization of EHV1 unprimed ponies with TK– EHV1 by intramuscular or intravenous injection did not induce any sign of disease occurrence. However, when vaccinated ponies were challenged with EHV1 virus, an anamnestic VN antibody response was observed. The ponies immunized with TK– EHV1 vaccine also showed less clinical signs of disease and live virus shedding to environment (Cornick et. al., 1990). However, live attenuated vaccine for EHV1 may not always produce optimal immune response. The intranasal administration of a live attenuated EHV1 vaccine (Rhinomune, Pfizer) failed to induce mucosal antibodies, which could explain the failure of attenuated EHV1 vaccines to prevent virus shedding. It was speculated that the attenuation may have reduced the virus’s mucosal immunogenicity (Breathnach et. al., 2001).
It was observed that mutant EHV1 viruses deficient in the whole IR (vL11ΔIR) produced plaques of small size and showed belated growth kinetics (Ahn et. al., 2011). The CBA mice intranasally infected demonstrated increased survival rate and decreased viral titre in the lungs compared to mice infected with parental or revertant virus.
Poxvirus–Based Vector Vaccine for EHV1
In live vector vaccine the epitope specific gene of pathogens are inserted in to genome of nonpathogenic infectious viruses through modern biotechnology tools. These viruses will express the epitope specific proteins in the host cell and will induce both humoral as well as cell mediated immune response. For such vaccine preparation recombinant poxviruses are generally used (Moss, 1996) because they are genetically stable and can accommodate relatively larger size inserts.
Several recombinant vaccines for horses have been developed from recombinant pox viruses such as vaccinia virus and avipoxvirus (Minke et. al., 2006). A new recombinant vaccinia virus derived (NYVAC strain) vaccine specific to IE protein (CTL target protein), has been introduced for vaccination in horses. The EHV1 vaccines derived from vaccinia virus can stimulate CMI in the horse and therefore induce protective immunity against the disease (Paillot et. al., 2006).
DIAGNOSIS
A number of diagnostic techniques based on microbiology and modern molecular biology tools have been used for diagnosis of EHV1. However, the ‘gold standard’ test for EHV1 infection diagnosis remains virus isolation. For estimation of EHV5 load in lung, the broncho–alveolar lavage is considered as good clinical sample (Marenzoni et. al., 2011). The EHV1 specific antigen and nucleic acid was detected in the trophoblasts of fetal membranes in spontaneous cases of equine abortion (Szerdi et. al., 2003).
The serum neutralization (SN) assay is traditional method for detection of EHV1 antibodies. However, it is unable to accurately differentiate the infected animals from vaccinated animals. However, in some experimental studies, co–relation of protection with SN titers has been reported (Goehring et. al., 2010; Goodman et. al., 2006). A further difficulty is the cross–reaction of EHV1 and EHV4 in SN assay (Hartley et. al., 2005).
A number of PCR based assays have been developed to target different genes of EHV1 such as thymidine kinase (TK) gene (Carvalho et. al.,2000) and various glycoprotein genes such as B (Borchers and Slater, 1993), C (Galosi et. al.,2001), D (Galosi et. al.,2001) and H (Varrasso et. al.,2001) for correct diagnosis. However, these techniques have some serious limitations in terms of being cumbersome and carry increased risk of cross contamination resulting from manipulation of amplified PCR product. However, few of these assays are either semi–nested or nested PCRs or used in combination with Southern blotting technique (Borchers and Slater, 1993; O’Keefe et. al., 1991).
Now–a–days, the real–time PCR has become a very valuable diagnostic tool in virology (Mackay et. al., 2002). The technique utilizes the combined activity of specific hybridization of a fluorogenic probe to a target gene and 5’ exonuclease activity of Taq DNA polymerase. The target nucleic acid sequence is amplified and simultaneously Taq hydrolyses the bound dual–labeled probe. This in turn shows fluorescence which is detected and recorded by software and plotted as a graph (Livak et. al., 1995; Heid et. al., 1996). The real–time PCR assay is very specific, fast, sensitive, and effective in diagnosis of viral diseases. It also has the advantage of low risk of cross–contamination over conventional PCR.
The glycoprotein B gene specific multiplex real–time PCR has the potential to distinguish EHV1 and EHV4 (Diallo et. al., 2007). To each equine herpes virus, specific primers and probes were designed and used in monoplex or multiplex PCR. The EHV1 and EHV4, specific minor–groove binding probes (MGBTM) labelled with 6–carboxyfluorescein (FAMTM) and VICR probes respectively are commonly used for detection.
A real–time PCR assay using allelic discrimination (E2) to distinguish between neuropathogenic and non–neuropathogenic strains of EHV1 has been reported (Allen, 2007). It was observed that the neuropathogenic phenotype of EHV1 was associated with a single–nucleotide polymorphism (A2254 or G2254) in open reading frame 30 (ORF30). Undoubtly this real– time PCR assay can differentiate neuropathogenic and non–neuropathogenic strains. However, it lacks sensitivity for routine viral nucleic acid detection from various clinical specimens. Similarly, the development of ORF30 region specific, allelic discrimination (E1) EHV1 real–time PCR test has been reported (Smith et. al., 2012).
Moreover, nasal swabs and blood samples used for rapid quantitative PCR testing have become standard practice for the diagnosis of active infections of EHV1 (Perkins et. al., 2008).
CONCLUSION AND FUTURE DIRECTIONS
Among the nine equine herpes virus types isolated from equines, the equine herpesvirus 1 (EHV1) and equine herpes virus 4 (EHV4) are responsible for significant economic losses to the equine industry globally. The infection with EHV1 can cause abortion, neonatal foal mortality, paralysis, and respiratory disease; while respiratory disease is the predominant outcome of EHV4 infection. The incidence of EHV1 induced equine herpesvirus myeloencephalopathy (EHM) has increased throughout the world during last decade. A single nucleotide polymorphism in EHV1 polymerase gene has been correlated with EHM, which may provide a selective advantage to neuropathogenic strain. The identification of factors playing a role in possible emergence of neuropathogenic strain needs to be investigated. The other reason of EHM could be reactivation/ recrudescence of latent EHV1 infection. Latency and reactivation are the key features of EHV infection which indicates the survival strategy of virus during course of evolution. However, our knowledge about EHV latency is quite scanty and it remains an underexplored area. Isolation of equine herpes viruses from new host species and new locations indicates the dynamic epidemiological picture of EHVs. The role of cell–associated viremia, latency and immune evasion in pathogenesis and epidemiology need further studies which in turn will help in devising control strategies.
Equine herpesvirus infection is difficult to clinically differentiate from other causes of equine respiratory disease, such as equine influenza, solely on the basis of clinical signs. Definitive diagnosis is determined by PCR or virus isolation from blood and the samples obtained from nose and throat. Suspected cases of EHV1 abortion can be confirmed by virus isolation, PCR and characteristic gross and microscopic lesions in the aborted foetus. Diagnostic techniques such as immunohistochemistry and PCR have diagnosed the cases of foetus negative, placenta–positive EHV associated abortion. Serological testing of mares after abortion shows little diagnostic value. Real–time PCR has shown its usefulness in diagnosing suspected cases of EHM. Also, there is need of a faster test to distinguish between the neurological strain and the non–neurological strain.
Gaps exist in our understanding of the correlates of protective immunity against EHV1. Vaccination against EHV is widely used especially among racehorses and brood mares, which helps in reducing virus shedding and duration of viraemia. None of current commercial vaccines is able to consistently prevent infection in a vaccinated animal or provide complete protection against diseases associated with EHV1. Currently used EHV vaccines have been shown to decrease clinical signs only in respiratory and abortive form of EHV infection. None of the current commercial vaccines gives protection against neurological form of the disease. There is a need to develop effective EHV1 vaccines that induce both mucosal and systemic cellular as well as humoral immunity in the horses along with a safe and efficient delivery route.
REFERENCES
Ahn BC, Zhang Y, Osterrieder N, O'Callaghan DJ (2011). Properties of an equine herpesvirus 1 mutant devoid of the internal inverted repeat sequence of the genomic short region. Virology. 410 (2): 327 – 335.
http://dx.doi.org/10.1016/j.virol.2010.11.020
PMid:21176938 PMCid:PMC3030640
Albrecht RA, Kim SK, O'Callaghan DJ (2005). The EICP27 protein of equine herpesvirus 1 is recruited to viral promoters by its interaction with the immediate–early protein. Virology. 333(1): 74 – 87.
http://dx.doi.org/10.1016/j.virol.2004.12.014
PMid:15708594
Allen GP, Bryans JT (1986). Molecular epizootiology, pathogenesis and prophylaxis of equine herpesvirus 1 infections. Prog. Vet. Microbiol. Immunol. 2: 78 – 144.
PMid:2856183
Allen GP, Kydd JH, Slater JD, Smith KC (1998). Advances in understanding of the pathogenesis, epidemiology and immunological control of equine herpesvirus abortion. Proceedings 8th International Conference on Equine Infectious Diseases, Dubai, 129 – 146.
PMid:9586775
Allen GP (2007). Development of a real–time polymerase chain reaction assay for rapid diagnosis of neuropathogenic strains of equine herpesvirus–1. J. Vet. Diagn. Invest. 19: 69 – 72.
http://dx.doi.org/10.1177/104063870701900110
PMid:17459834
Allen GP, Bolin DC, Bryant U, Carter CN, Giles RC, Harrison LR, Hong CB, Jackson CB, Poonacha K, Wharton R, Williams NM (2008). Prevalence of latent, neuropathogenic equine herpesvirus–1 in the thoroughbred broodmare population of central Kentucky. Equine Vet. J. 40(2): 105 – 110.
http://dx.doi.org/10.2746/042516408X253127
PMid:18089469
Annelies P, Vandekerckhove S, Glorieux AC, Gryspeerdt L, Steukers L, Duchateau N, Osterrieder GR, Van de Walle, Nauwynck HJ (2010). Replication kinetics of neurovirulent versus non–neurovirulent equine herpesvirus type 1 strains in equine nasal mucosal explants. J. Gen. Virol. 91: 2019 – 2028.
http://dx.doi.org/10.1099/vir.0.019257-0
PMid:20427565
Awan AR, Gibson JS, Field HJ (1991). A murine model for studying EHV1–induced abortion. Res. Vet. Sci. 51: 94 – 99.
http://dx.doi.org/10.1016/0034-5288(91)90038-P
Azab W, Osterrieder N (2012). Type 1 (EHV–1) and EHV–4 determine glycoproteins D of equine herpesvirus cellular tropism independently of integrins. J. Virol. 86(4): 2031 – 2044.
http://dx.doi.org/10.1128/JVI.06555-11
PMid:22171258 PMCid:PMC3302398
Barrandeguy M, Vissani A, Olguin C, Becerra L, Mi-o S, Pereda A, Oriol J, Thiry E (2008). Experimental reactivation of equine herpesvirus–3 following corticosteroid treatment. Equine Vet. J. 40(6): 593 – 595.
http://dx.doi.org/10.2746/042516408X333399
PMid:19031515
Baxi MK, Efstathiou S, Lawrence G, Whalley JM, Slater JD, Field HJ (1995). The detection of latency associated transcripts of equine herpesvirus1 in ganglionic neurons. J. Gen. Virol. 76(12): 3113 – 3118.
http://dx.doi.org/10.1099/0022-1317-76-12-3113
PMid:8847517
Belák S, Pàlfi V, Tuboly S, Bartha L (1980). Passive immunization of foals to prevent respiratory disease caused by equine herpesvirus type 2. Zentralbl. Veterinarmed. B. 27: 826 – 830.
http://dx.doi.org/10.1111/j.1439-0450.1980.tb02037.x
PMid:6164185
Borchers K, Slater J (1993). A nested PCR for the detection and differentiation of EHV–1 and EHV–4. J. Vir. Meth. 45: 331 – 336.
http://dx.doi.org/10.1016/0166-0934(93)90117-A
Borchers K, Wolfinger U, Ludwig H (1999). Latency–associated transcripts of equine herpesvirus type 4 in trigeminal ganglia of naturally infected horses. J. Gen. Virol. 80(8): 2165 – 2171.
PMid:10466816
Breathnach CC, Yeargan MR, Sheoran AS, Allen JP (2001). The mucosal humoral immune response of the horse to infective challenge and vaccination with equine herpesvirus–1 antigens. Equine Vet. J. 33(7): 651 – 657.
http://dx.doi.org/10.2746/042516401776249318
PMid:11770985
Brosnahan MM, Osterrieder N (2009). Equine herpesvirus–1: a review and update. In: T. S. Mair and R. E. Hutchinson (eds), Infectious Diseases of the Horse, Equine Veterinary Journal Ltd., Fordham, 41–51.
Browning GF, Bulach DM, Ficorelli N, Roy EA, Thorp BH, Studdert MJ (1988). Latency of equine herpes virus 4 (equine rhinopneumonitis). Vet. Rec. 123: 518 – 519.
http://dx.doi.org/10.1136/vr.123.20.518
PMid:2849833
Bryans JT, Prickett ME (1969). A consideration of the pathogenesis of abortigenic disease caused by equine herpesvirus 1. Proceedings 2nd International Conference on Equine Infectious Diseases, 34 – 40.
Bungener L, Huckriede A, Wilschut J, Deamen T (2002). Delivery of protein antigens to the immune system by fusion–active virosomes: a comparison with liposomes and ISCOMs. Biosci. Rep. 22(2): 323 – 338.
http://dx.doi.org/10.1023/A:1020198908574
PMid:12428908
Carvalho R, Passos LMF, Martins S (2000). Development of a differential multiplex PCR assay for equine herpesvirus 1 and 4 as a diagnostic tool. J. Vet. Med. B. 47: 351 – 359.
http://dx.doi.org/10.1046/j.1439-0450.2000.00361.x
Caughman GB, Staczek J, O'Callaghan DJ (1985). Equine herpesvirus type 1 infected cell polypeptides: evidence for immediate early/early/late regulation of viral gene expression. Virology. 145: 49 – 61.
http://dx.doi.org/10.1016/0042-6822(85)90200-4
Chester PM, Allsop R, Purewal A, Edington N (2007). Detection of latency–associated transcripts of equid herpesvirus 1 in equine leucocyte but not in trigeminal ganglia. J. Virol. 71(5): 3437 – 3443.
Cook RF, O'Neill T, Strachan E, Sundquist B, Mumford JA (1990). Protection against lethal equine herpes virus type 1 (subtype 1) infection in hamsters by immune stimulating complexes (ISCOMs) containing the major viral glycoproteins. Vaccine. 8(5): 491 – 496.
http://dx.doi.org/10.1016/0264-410X(90)90252-H
Cornick J, Martens J, Crandel R, McConnell S, Kit S (1990). Safety and efficacy of a thymidine kinase negative equine herpesvirus–1 vaccine in young horses. Can. J. Vet. Res. 54(2): 260 – 266.
PMid:2162730 PMCid:PMC1255647
Crabb BS, Studdert MJ (1996). Rhinopneumonitis (equine herpesvirus 4) and equine abortion (equine herpesvirus 1), in virus infections of equines. In: M.J. Studdert (ed). Virus Infections of Vertebrates, vol. 6. Elsevier, Amsterdam, (Chapter 2), 11 – 37.
Dargan DJ, Patel AH, Subak–Sharpe JH (1995). PREPs: herpes simplex virus type 1–specific particles produced by infected cells when viral DNA replication is blocked. J. Virol. 69(8): 4924 – 4932.
PMid:7609061 PMCid:PMC189307
Dargan DJ, Subak–Sharpe JH (1997). The effect of herpes simplex virus type 1 L–particles on virus entry, replication, and the infectivity of naked herpesvirus DNA. Virology. 239(2): 378 – 388.
http://dx.doi.org/10.1006/viro.1997.8893
PMid:9434728
Darlington RW, Randall CC (1963): The nucleic acid content of equine abortion virus. Virology. 19: 322 – 327.
http://dx.doi.org/10.1016/0042-6822(63)90071-0
Davison AJ (2002). Evolution of the herpesviruses. Vet. Microbiol. 86(1–2): 69 – 88.
http://dx.doi.org/10.1016/S0378-1135(01)00492-8
Davison AJ, Eberle R, Ehlers B, Hayward GS, McGeoch DJ, Minson AC, Pellett PE, Roizman B, Studdert MJ, Thiry E (2009). The order Herpesvirales. Arch. Virol. 154: 171 – 177.
http://dx.doi.org/10.1007/s00705-008-0278-4
PMid:19066710 PMCid:PMC3552636
Diallo IS, Hewitson G, Wright L, Kelly MA, Rodwell BJ, Corney BG (2007). Multiplex real–time PCR for the detection and differentiation of equid herpesvirus 1 (EHV–1) and equid herpesvirus 4 (EHV–4). Vet. Microbiol. 123: 93 – 103.
http://dx.doi.org/10.1016/j.vetmic.2007.02.004
PMid:17346907
Edington N, Bridges CG, Huckle A (1985). Experimental reactivation of equid herpesvirus 1 (EHV–1) following administration of corticosteroids. Equine Vet. J. 17: 369 – 372.
http://dx.doi.org/10.1111/j.2042-3306.1985.tb02524.x
PMid:2996879
Edington N, Bridges CG, Patel JR (1986). Endothelial cell infection and thrombosis in paralysis caused by equid herpesvirus–1: equine stroke. Arch. Virol. 90: 111 – 124.
http://dx.doi.org/10.1007/BF01314149
PMid:3015074
Edington N, Smyth B, Griffiths L (1991). The role of endothelial cell infection in the endometrium, placenta and foetus of equid herpesvirus–1 (EHV–1) abortions. J. Comp. Path. 104: 379 – 387.
http://dx.doi.org/10.1016/S0021-9975(08)80148-X
Edington N, Chesters P, Azam S, Welch H, McGladdery A, Purewal A (1994). Profiles of a herpesviruses in circulating leucocytes from thoroughbred mares and foals using PCR and co–cultivation. Equine Infect. Dis.7: 251 – 254.
El–Habashi N, Kato Y, EL–Nahass E, Fukushi H, Hirata A, Sakai H, Kimura J, Yanai T (2013). An ocular infection model using suckling hamsters inoculated with equine herpesvirus 9 (EHV–9): kinetics of the virus and time–course pathogenesis of EHV–9–induced encephalitis via the eyes. Vet. Pathol. 50 (1): 56 – 6.
http://dx.doi.org/10.1177/0300985812442691
PMid:22508701
Foote CE, Love DN, Gilkerson JR, Rota J, Trevor–Jones P, Ruitenberg KM, Wellington JE, Whalley JM (2005). Serum antibody responses to equine herpesvirus 1 glycoprotein D in horses, pregnant mares and young foals. Vet. Immunol. Immunopathol. 105(1–2): 47 – 57.
http://dx.doi.org/10.1016/j.vetimm.2004.12.012
PMid:15797474
Foote CE, Raidal SL, Pecenpetelovska G, Wellington JE, Whalley JM (2006). Inoculation of mares and very young foals with EHV–1 glycoproteins D and B reduces virus shedding following respiratory challenge with EHV–1. Vet. Immunol. Immunopathol. 111(1–2): 97 – 108.
http://dx.doi.org/10.1016/j.vetimm.2006.01.012
PMid:16504306
Frampton AR, Uchida H, Einem JV, Goins WF, Grandi P, Cohen JB, Osterrieder N, Glorioso JC (2010). Equine herpesvirus type 1 (EHVa H, Einem JV, Goins WF, Grandi P, Cohen JB, Osterrieder N, Glorioso JC (2010). Equine herpesvirus type 1 (EHV–1) utilizes microtubules, dynein, and ROCK1 to productively infect cells. Vet. Microbiol. 141(1–2): 12 – 21.
http://dx.doi.org/10.1016/j.vetmic.2009.07.035
PMid:19713056 PMCid:PMC2819619
Fuentealba N, Sguazza G, Scrochi M, Bravi M, Zanuzzi C, Corva S, Gimeno E, Pecoraro M, Galosi C (2014). Production of equine herpesvirus 1 recombinant glycoprotein D and development of an agar gel immunodiffusion test for serological diagnosis. J. Virol. Methods. 202: 15 – 18.
http://dx.doi.org/10.1016/j.jviromet.2014.02.025
PMid:24631347
Galosi CM, Vila Roza MV, Oliva GA, Pecorado MR, Echeverria MG, Corva S, Etcheverrigaray ME (2001). A polymerase chain reaction for detection of equine herpesvirus–1 in routine diagnostic submissions of tissues from aborted fetuses. J. Vet. Med. 48: 341 – 346.
http://dx.doi.org/10.1046/j.1439-0450.2001.00455.x
Goehring LS, van Maanen C, Sloet van Oldruitenborgh–Oosterbaan MM (2005). Neurological syndromes among horses in the Netherlands: a 5 year retrospective survey (1999–2004). Vet. Q. 27: 11 – 20.
http://dx.doi.org/10.1046/j.1439-0450.2001.00455.x
PMid:15835280
Goehring LS, Wagner B, Bigbie R, Hussey SB, Rao S, Morley PS, Lunn DP (2010). Control of EHV1 viremia and nasal shedding by commercial vaccines. Vaccine. 28(32): 5203 – 5211.
http://dx.doi.org/10.1016/j.vaccine.2010.05.065
PMid:20538091
Goodman LB, Wagner B, Flaminio MJ, Sussman KH, Metzger SM, Holland R, Osterrieder N (2006). Comparison of the efficacy of inactivated combination and modified–live virus vaccines against challenge infection with neuropathogenic equine herpesvirus type 1 (EHV–1). Vaccine. 24: 3636 – 3645.
http://dx.doi.org/10.1016/j.vaccine.2006.01.062
PMid:16513225
Hartley CA, Wilks CR, Studdert MJ, Gilkerson JR (2005). Comparison of antibody detection assays for the diagnosis of equine herpesvirus 1 and 4 infections in horses. Am. J. Vet. Res. 66: 921 – 928.
http://dx.doi.org/10.2460/ajvr.2005.66.921
PMid:15934623
Hasebe R, Sasaki M, Sawa H, Wada R, Umemura T, Kimura T (2009). Infectious entry of equine herpesvirus–1 into host cells through different endocytic pathways. Virology. 393(2): 198 – 209.
http://dx.doi.org/10.1016/j.virol.2009.07.032
PMid:19720389
Hebia I, Fieni F, Duchamp G, Destrumelle S, Pellerin JL, Zientara S, Vautherot JF, Bruyas JF (2007). Potential risk of equine herpes virus 1 (EHV–1) transmission by equine embryo transfer. Theriogenology. 67(9): 1485 – 1491.
http://dx.doi.org/10.1016/j.theriogenology.2007.03.006
PMid:17459463
Heid CA, Stevens J, Livak KJ, Williams PM (1996). Real–time quantitative PCR. Genome. Res. 6: 986 – 994.
http://dx.doi.org/10.1101/gr.6.10.986
PMid:8908518
Henninger RW, Reed SM, Saville WJ, Allen GP, Hass GF, Kohn CW, Sofaly C (2007). Outbreak of neurologic disease caused by equine herpesvirus–1 at a university equestrian center. J. Vet. Intern. Med. 21: 157 – 165.
http://dx.doi.org/10.1111/j.1939-1676.2007.tb02942.x
PMid:17338164
Jackson TA, Osburn BI, Cordy DR, Kendrick JW (1977). Equine herpesvirus 1 infection of horses: studies on the experimentally induced neurologic disease. Am. J. Vet. Res. 38: 709 – 719.
PMid:195494
Kapoor S, Dhama K, Pawaiya RVS, Mathew T (2010). Equine Herpesvirus. In: Tropical Viral Diseases of Large Animals Part 1. Editor, Thankam Mathew. Published by Thajema Publishers, West Orange, New Jersey, USA. Pp110 – 132.
PMid:20331828
Kukreja A, Walker C, Fitzmaurice T, Awan AR, Love DN, Whalley JM, Field HJ (1998). Protective effects of equine herpesvirus–1 (EHV–1) glycoprotein B in a murine model of EHV–1–induced abortion. Vet. Microbiol. 62(4): 303 – 311.
http://dx.doi.org/10.1016/S0378-1135(98)00211-9
Livak KJ, Flood SJ, Marmaro J, Giusti W, Deetz K (1995). Oligonucleotides with fluorescent dyes at opposite ends provide a quenched system useful for detecting PCR product and nucleic acid hybridisation. PCR Meth. 4: 357 – 362.
http://dx.doi.org/10.1101/gr.4.6.357
Lunn DP, Davis–Poynter N, Flaminio MJ, Horohov DW, Osterrieder K, Pusterla N, Townsend HG (2009). Equine herpesvirus–1 consensus statement. J. Vet. Intern. Med. 23: 450 – 461.
http://dx.doi.org/10.1111/j.1939-1676.2009.0304.x
PMid:19645832
Mackay IM, Arden KE, Nitsche A (2002). Survey and summary: real time in virology. Nucl. Acids. Res. 30: 1292 – 1305.
http://dx.doi.org/10.1093/nar/30.6.1292
PMid:11884626 PMCid:PMC101343
Marenzoni ML, Passamonti F, Lepri E, Cercone M, Capomaccio S, Cappelli K, Felicetti M, Coppola G, Coletti M, Thiry E (2011). Quantification of Equid herpesvirus 5 DNA in clinical and necropsy specimens collected from a horse with equine multinodular pulmonary fibrosis. J. Vet. Diagn. Invest. 23 (4): 802 – 806.
http://dx.doi.org/10.1177/1040638711407890
PMid:21908328
Marshall KR, Field HJ (1997). Demonstration of equine herpesvirus–1 neuronal latency in murine olfactory bulbs using a novel combined in situ PCR and protein synthesis method. Virology. 229: 279 – 282.
http://dx.doi.org/10.1006/viro.1996.8394
PMid:9123871
Mc Geoch DJ, Dolan A, Ralph AC (2000). Toward a comprehensive phylogeny for mammalian and avian herpesviruses. J. Virol. 74(22): 10401 – 10406.
http://dx.doi.org/10.1128/JVI.74.22.10401-10406.2000
McLauchlan J, Rixon FJ (1992). Characterization of enveloped tegument structures (L particles) produced by alphaherpesviruses: integrity of the tegument does not depend on the presence of capsid or envelope. J. Gen. Virol. 73 (2): 269 – 276.
http://dx.doi.org/10.1099/0022-1317-73-2-269
PMid:1311356
Minke JM, Fischer L, Baudu P, Guigal PM, Sindle T, Mumford JA, Audonnet JC (2006). Use of DNA and recombinant canarypox viral (ALVAC) vectors for equine herpes virus vaccination. Vet. Immunol. Immunopathol. 111(1–2): 47 – 57.
http://dx.doi.org/10.1016/j.vetimm.2006.01.008
PMid:16580075
Mori CMC, Mori E, Favaro LL, Santos CR, Lara MCCSH, Villalobos EMC, Cunha EMS, Brandao PE, Richtzenhain LJ, Maiorka PC (2012). Equid herpesvirus type–1 exhibits neurotropism and neurovirulence in a mouse model. J. Comp. Path. 146: 202 – 210.
http://dx.doi.org/10.1016/j.jcpa.2011.04.003
PMid:21689826
Moss B (1996). Genetically engineered poxviruses for recombinant gene expression, vaccination, and safety. Proc. Natl. Acad. Sci. U S A. 93(21): 11341 – 11348.
http://dx.doi.org/10.1073/pnas.93.21.11341
PMid:8876137 PMCid:PMC38059
Mumford JA, Hannant D, Jesset DM, O'Neill T, Smith KC, Ostlund E (1994). Abortigenic and neurological disease caused by experimental infection with equid herpesvirus–1. Proceedings 7th International Conference on Equine Infectious Diseases, Tokyo. 261 – 275.
Nageshi HS, McNeil JR, Ficorilli N, Studdert MJ (1992). Cloning and restriction endonuclease mapping of the genome of an equine herpesvirus 4 (equine rhinopneumonitis virus), strain 405/76. Arch. Virol. 124: 379 – 387.
http://dx.doi.org/10.1007/BF01309818
Nowotny N, Burtscher H, Biirki F (1987). Neuropathogenicity for suckling mice from the lippizan outbreak 1983 and of selected other EHV1 strains. J. Vet. Med. B. 34: 441 – 448.
http://dx.doi.org/10.1111/j.1439-0450.1987.tb00418.x
Nugent J, Birch–Machin I, Smith KC, Mumford JA, Swann Z, Newton JR, Bowden RJ, Allen GP, Davis–Poynter N (2006). Analysis of equid herpesvirus 1 strain variation reveals a point mutation of the DNA polymerase strongly associated with neurovirulent versus nonneurovirulent disease outbreaks. J. Virol. 80. 4047 – 4060.
http://dx.doi.org/10.1128/JVI.80.8.4047-4060.2006
PMid:16571821 PMCid:PMC1440451
O'Keefe JS, Murray A, Wilks CR, Moriarty KM (1991). Amplification and differentiation of the DNA of an abortigenic (type 1) and a respiratory (type 4) strain of equine herpesvirus by polymerase chain reaction. Res. Vet. Sci. 50: 349 – 351.
http://dx.doi.org/10.1016/0034-5288(91)90137-D
Paillot R, Ellis SA, Daly JM, Audonnet JC, Minke JM, Davis–Poynter N, Hannant D, Kydd JH (2006). Characterisation of CTL and IFN–γ synthesis in ponies following vaccination with a NYVAC based construct coding for EHV–1 immediate early gene, followed by challenge infection. Vaccine. 24(10): 1490 – 1500.
http://dx.doi.org/10.1016/j.vaccine.2005.10.019
PMid:16269205
Pardoe I, Dargan D (2002). PREPS and L–particles: a new approach to virus–like particle vaccines. Expert Rev. Vaccines. 1(4): 427 – 432
http://dx.doi.org/10.1586/14760584.1.4.427
PMid:12901580
Patel JR, Edington N, Mumford JA (1982). Variation in cellular tropism between isolates of equine herpesvirus–1 in foals. Arch. Virol. 74: 41 – 51.
http://dx.doi.org/10.1007/BF01320781
PMid:6297429
Patel JR, Heldens J (2005). Equine herpesviruses 1 (EHV–1) and 4 (EHV–4) – epidemiology, disease and immunoprophylaxis: a brief review. Vet. J. 170: 14 – 23.
http://dx.doi.org/10.1016/j.tvjl.2004.04.018
PMid:15993786
Perkins G, Ainsworth DM, Erb HN, Del Piero F, Miller M, Wilkins PA, Palmer J, Frazer M (1999). Clinical, haematological and biochemical findings in foals with neonatal equine herpesvirus–1 infection compared with septic and premature foals. Equine Vet. J. 31: 422 – 426.
http://dx.doi.org/10.1111/j.2042-3306.1999.tb03843.x
PMid:10505959
Perkins GA, Goodman LB, Dubovi EJ, Kim SG, Osterrieder N (2008). Detection of equine herpesvirus–1 in nasal swabs of horses by quantitative real–time PCR. J. Vet. Intern. Med. 22: 1234 – 1238.
http://dx.doi.org/10.1111/j.1939-1676.2008.0172.x
PMid:18691363
Plummer G, Goodheart CR, Studdert MJ (1973). Equine herpesviruses: antigenic relationships and deoxyribonucleic acid densities. Infect. Immu. 8: 621 – 627.
PMid:4742974 PMCid:PMC422900
Prickett ME (1969).The pathology of disease caused by equine herpesvirus 1. Proceedings 2nd International Conference on Equine Infectious Diseases. 24 – 33.
Pronost S, Léon A, Legrand L, Fortier C, Miszczak F, Freymuth F, Fortier G (2010). Neuropathogenic and non–neuropathogenic variants of equine herpesvirus 1 in France. Vet. Microbiol. 145(3–4): 329 – 333.
http://dx.doi.org/10.1016/j.vetmic.2010.03.031
PMid:20427133
Pusterla N, Wilson WD, Mapes S, Finno C, Isbell D, Arthur RM, Ferraro GL (2009). Characterization of viral loads, strains and state of equine herpesvirus–1 using real–time PCR in horses following natural exposure at a racetrack in California. Vet. J. 179(2): 230 – 239.
http://dx.doi.org/10.1016/j.tvjl.2007.09.018
PMid:18024200
Pusterla N, Mapes S, Wilson WD (2012). Prevalence of latent alpha herpesviruses in thoroughbred racing horses. Vet. J. 193(2): 579 – 582.
http://dx.doi.org/10.1016/j.tvjl.2012.01.030
PMid:22405721
Roizman B, Desrosiers RS, Fleckenstein B, Lopez C, Minson MJ, Studdert MJ (1992). The family Herpesviridae: an update. Arch. Virol. 123: 425 – 449.
http://dx.doi.org/10.1007/BF01317276
Roizman B (1996). Herpesviridae.In: B.N. Field, D.M. Knipe, P.M. Howley, R.M. Channock, J.L. Melnick, T.P. Monath, B. Roizman, S.E. Straus (eds), Virology, third ed. Lippincott–Raven, Philadelphia, PA, 2221 – 2230.
Ruitenberg KM, Walker C, Wellington JE, Love DN, Whalley JM (1999). DNA–mediated immunization with glycoprotein D of equine herpesvirus 1 (EHV– 1) in a murine model of EHV–1 respiratory infection. Vaccine. 17(3): 237 – 244.
http://dx.doi.org/10.1016/S0264-410X(98)00192-3
Ruitenberg KM, Love DN, Gilkerson JR, Wellington JE, Whalley JM (2000). Equine herpesvirus 1 (EHV–1) glycoprotein D DNA inoculation in horses with preexisting EHV–1/EHV–4 antibody. Vet. Microbiol. 76(2): 117 – 127.
http://dx.doi.org/10.1016/S0378-1135(00)00237-6
Sabine M, Robertson GR, Whalley JM (1981). Differentiation of sub–types of equine herpesvirus 1 by restriction endonuclease analysis. Aust. Vet. J. 57: 148 – 149.
http://dx.doi.org/10.1111/j.1751-0813.1981.tb00495.x
PMid:6266385
Slater J (2007). Equine herpesviruses. In: D.C. Sellon, M.T. Long (eds), Equine Infectious Diseases. St. Louis: Saunders Elsevier. 134 – 152.
http://dx.doi.org/10.1016/B978-1-4160-2406-4.50018-1
Smith KC, Whitwell KE, Binns MM, Dolby CA, Hannant D, Mumford JA (1992). Abortion of virologically negative foetuses following experimental challenge of pregnant pony mares with equid herpesvirus 1. Equine Vet. J. 24: 256 – 259.
http://dx.doi.org/10.1111/j.2042-3306.1992.tb02830.x
PMid:1323457
Smith KL, Li Y, Breheny P, Cook RF, Henney PJ, Sells S, Pronost S, Lu Z, Crossley BM, Timoney PJ, Balasuriya UBR (2012). New real–time PCR assay using allelic discrimination for detection and differentiation of equine herpesvirus–1 strains with A2254 and G2254 polymorphisms. J. Clin. Microbiol. 50 (6): 1981 – 1988
http://dx.doi.org/10.1128/JCM.00135-12
PMid:22493339 PMCid:PMC3372139
Smith PM, Zhang Y, Grafton WD, Jennings SR, O'Callaghan DJ (2000). Severe murine lung immunopathology elicited by the pathogenic equine herpesvirus 1 strain RacL11 correlates with early production of macrophage inflammatory proteins lalpha, lbeta, and 2 and tumor necrosis factor alpha. J. Virol. 74: 1034 – 1040.
http://dx.doi.org/10.1128/JVI.74.21.10034-10040.2000
Stevens JG, Wagner EK, Devi–Rao GB, Cook ML, Feldman LT (1987). RNA complimentary to a herpesvirus alpha gene mRNA is prominent in latently infected neurons. Sci. 235: 1056 – 1059.
http://dx.doi.org/10.1126/science.2434993
Stevens JG (1989). Human herpesvirus: a consideration of the latent state. Microbiol. Rev. 53: 318 – 332.
PMid:2552271 PMCid:PMC372739
Stevens JG (1994). Overview of herpesvirus latency. Semin. Virol. 5:191 – 196.
http://dx.doi.org/10.1006/smvy.1994.1020
Stroop WG, Rock DL, Fraser NW (1984). Localization of herpes simplex virus in the trigeminal and olfactory systems of the mouse central nervous system during acute and latent infections by in situ hybridization. Lab. Invest. 51: 27 – 38.
PMid:6330452
Studdert MJ, Simpson T, Roizman B (1981). Differentiation of respiratory and abortogenic isolates of equine herpesvirus 1 by restriction endonucleases. Sci. 214: 562 – 564.
http://dx.doi.org/10.1126/science.6270790
Studdert MJ, Hartley CA, Dynon K, Sandy JR, Slocombe RF, Charles JA, Milne ME, Clarke AF, El Hage C (2003). Outbreak of equine herpesvirus type 1 myeloencephalitis: new insights from virus identification by PCR and the application of an EHV–1–specific antibody detection ELISA. Vet. Rec. 153: 417 – 423.
http://dx.doi.org/10.1136/vr.153.14.417
PMid:14582730
Szeredi L, Aupperle H, Steiger K (2003). Detection of equine herpesvirus–1 in the fetal membranes of aborted equine fetuses by immunohistochemical and in–situ hybridization techniques. J. Comp. Path. 129: 147 – 153.
http://dx.doi.org/10.1016/S0021-9975(03)00022-7
Teleford EAR, Watson MS, McBride K, Davison AJ (1992). The DNA sequence of equine herpesvirus–1. Virol. 189: 304 – 316.
http://dx.doi.org/10.1016/0042-6822(92)90706-U
Teleford EAR, Watson MS, Perry J, Cullinane AA, Davison AL (1998). The DNA sequence of equine herpesvirus–4. J. Gen. Virol. 79: 1197 – 1203.
Tewari D, Nair SV, De Ungria MC, Lawrence GL, Hayden M, Love DN, Field HJ, Whalley JM (1995). Immunization with glycoprotein C of equine herpesvirus–1 is associated with accelerated virus clearance in a murine model. Arch. Virol. 140(4): 789 – 797.
http://dx.doi.org/10.1007/BF01309967
PMid:7794119
Thein P (1978). The association of EHV–2 infection with keratitis and research on the occurrence of equine coital exanthema (EHV–3) of horses in Germany. In: J.T. Bryans and H. Gerber (eds) Equine infectious disease IV. Princeton New Jersey: Veterinary Publications, 33 – 41.
Thein P, Darai G, Janssen W, Berg le RD, Strube W, Floss G (1993). Recent information about the etiopathogenesis of paretic–paralytic forms of herpesvirus infection in horses. Tierärztl. Prax. 21: 445 – 450.
PMid:8248905
van Maanen C, Willink DL, Smeenk LA, Brinkhof J, Terpstra C (2000). An equine herpesvirus 1 (EHV1) abortion storm at a riding school. Vet. Q. 22(2): 83 – 87.
http://dx.doi.org/10.1080/01652176.2000.9695030
PMid:10789515
Van de Walle Gerlinde R, Goupil R, Wishon C, Damiani A, Perkins GA, Osterrieder N (2009). A single–nucleotide polymorphism in a herpesvirus DNA polymerase is sufficient to cause lethal neurological disease. J. Infect. Dis. 200: 20 – 25.
http://dx.doi.org/10.1086/599316
PMid:19456260
Varrasso A, Dynon K, Ficorelli N, Hartley CA, Studdert MJ, Drummer HE (2001). Identification of equine herpesvirus 1 and 4 by polymerase chain reaction. Aust. Vet. J. 79: 563 – 569.
http://dx.doi.org/10.1111/j.1751-0813.2001.tb10751.x
PMid:11599819
Walker C, Love DN, Whalley JM (1999). Comparison of the pathogenesis of acute equine herpesvirus 1 (EHV–1) in the horse and the mouse model: a review. Vet. Microbiol. 68: 3 – 13.
http://dx.doi.org/10.1016/S0378-1135(99)00056-5
Weerasinghe CU, Learmonth GS, Gilkerson JR, Foote CE, Wellington JE, Whalley JM (2006). Equine herpesvirus 1 glycoprotein D expressed in E. coli provides partial protection against equine herpesvirus infection in mice and elicits virus–neutralizing antibodies in the horse. Vet. Immunol. Immunopathol. 111(1–2): 59 – 66.
http://dx.doi.org/10.1016/j.vetimm.2006.01.009
PMid:16473414
Welch HM, Bridges CG, Lyon AM, Griffiths L, Edington N (1992). Latent equid herpesviruses 1 and 4: detection and distinction using the polymerase chain reaction and co–cultivation from lymphoid tissues. J. Gen. Virol. 73: 261 – 268.
http://dx.doi.org/10.1099/0022-1317-73-2-261
PMid:1347078
Whitwell KE, Blunden AS (1992). Pathological findings in horses dying during an outbreak of the paralytic form of equid herpesvirus type 1 (EHV–1) infection. Equine Vet. J. 24: 13 – 19.
http://dx.doi.org/10.1111/j.2042-3306.1992.tb02771.x
PMid:1313358
Williams mail KJ, Robinson NE, Lim A, Brandenberger C, Maes R, Behan A, Bolin SR (2013). Experimental induction of pulmonary fibrosis in horses with the gammaherpesvirus equine herpesvirus 5. Plos One.8 (10): e77754
http://dx.doi.org/10.1371/journal.pone.0077754
PMid:24147074 PMCid:PMC3795644
Wilson WD (1997). Equine herpesvirus 1 myeloencephalopathy. Vet. Clin. North Am. Equine Pract. 13: 53 – 72
PMid:9106343


