
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 2 (2S): 23 – 26Special Issue–2 (2014) (Advances in Diagnosis and Control of Infectious Diseases of Animals)
An Alternative Amplification and Sequencing Approach for Genome Segment 2 of Bluetongue Virus
Arnab Ghosh1, Narender Singh Maan2, Aman Kumar1, Kanisht Batra1, Mahendra Singh1, Sunayna1, Sushila Maan1*
- Department of Animal Biotechnology, College of Veterinary Sciences, Lala Lajpat Rai University of Veterinary and Animal Sciences, Hisar, 125 004, Haryana, India
- Department of Animal Nutrition, Resource faculty Department of Animal Biotechnology, College of Veterinary Sciences, Lala Lajpat Rai University of Veterinary and Animal Sciences, Hisar, 125 004, Haryana, India
*Corresponding author:sushilamaan105@gmail.com
ARTICLE CITATION:
Ghosh A, Maan NS, Kumar A, Batra K, Singh M, Sunayna, Maan S (2014). An alternative amplification and sequencing approach for genome segment 2 of bluetongue virus. Adv. Anim. Vet. Sci. 2 (2S): 23 – 26.
Received: 2014–03–08, Revised: 2014–04–12, Accepted: 2014–04–13
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/journal.aavs/2014/2.2s.23.26
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Bluetongue (BT) is an infectious but non–contagious disease of domestic and wild ruminants and is transmitted via the bite of infected Culicoides midges. Bluetongue virus (BTV), the prototype species of the genus Orbivirus (family Reoviridae) contains double–stranded (ds) RNA genome. The dsRNA genome is arranged as ten linear segments which are packaged as exactly one copy of each within the icosahedral protein capsid of the virus particle. The genome segments encode 7 structural proteins (VP1–VP7) and four non–structural proteins (NS1–NS4). BTV exists as 26 distinct serotypes, as well as different topotypes and also has the potential to exchange genome segments. Any serotype of BTV has the potential to cause disease in ruminants with high rate of mortality and significant economic losses. For the genetic analysis of these viruses, an alternative method to already published Full–Length Amplification of cDNAs’ (FLAC) technique (Maan et al., 2007), has been developed for the amplification of genome segment 2 of BTVs. In this method genome segment 2 of BTV–9 (~2.9kb) was amplified in four different overlapping fragments using four pairs of amplification primers. Each fragment was then sequenced using at least four sequencing primers. Sequences from all four fragments were then assembled to generate a contiguous sequence. This amplification and sequencing strategy is particularly useful when the initial amount of RNA in the sample is too low (e.g. tissue or blood samples from field) to perform FLAC and/or cloning. Using this approach, sequence data can be obtained from a new sample within few hours.
INTRODUCTION
Bluetongue (BT) is a non contagious, infectious, arthropod borne disease of domestic and wild ruminants. Bluetongue virus (BTV) belongs from the genus Orbivirus in the family Reoviridae and spread to susceptible host by insect vectors (biting midges) of Culicoides genus (Mellor et al., 2009). There were 24 historical serotypes of this virus worldwide. However, two further BTV serotypes, BTV–25 (Toggenburg orbivirus, from Switzerland) and BTV–26 (from Kuwait) have recently been identified from goats and sheep, respectively (Maan et al., 2011). Bluetongue is endemic in India, although BT appears to have great impact on the livestock sector, with exotic breeds of sheep, including Rambouillet, Russian Merino, Southdown, Corriedale and Suffolk initially thought to be more susceptible to disease. Twenty–two serotypes have been reported from India, thirteen (BTV–1, –2, –3, –4, –8, –9, –10, –12, –16, –17, –18, –21, –23) of these on the basis of virus isolation and nine (BTV–5, –6, –7, , –11, –13, –14, –15, –19, –20) on the basis of neutralizing antibodies (Prasad et al., 2009; Maan et al., 2012; Rao et al., 2013b). BTV has 10 segments of dsRNA genome encoding 7 structural (VP1 to VP7) and four non structural proteins (NS1, NS2, NS3/3a, and NS4) (Belhouchet et al., 2011). The core of the icosahedral virion is made up of two major proteins (VP3 and VP7) enclosing three minor protein (VP1, VP4 and VP6), and 10 dsRNA segments (Verwoerd and Huismans, 1972). Core is surrounded by a diffused outer layer of two major proteins (VP2 and VP5). VP2 is the major serotype determining protein of BTV which is encoded by most variable genome segment 2. Sequencing of genome segment 2 of BTV is one of the methods for serotype determination of the virus (Maan et al., 2007; Rao et al., 2013a). Full–Length Amplification of cDNAs’ (FLAC) is a reliable method to determine the full length sequences of complete genome of these viruses (Maan et al., 2007). However, sequencing of complete segment 2 of BTV by FLAC and/or cloning when the initial sample concentration is too low can be problematic. Here, we describe an alternative amplification and sequencing approach for segment 2 genome of BTV from samples where initial viral RNA concentration is too low.
MATERIALS AND METHODS
Virus and Cell Culture
Bluetongue virus serotype 9 (BTV–9) an Indian strain was grown in BHK–21 cell line for five passages in Dulbecco’s minimum essential medium (DMEM) supplemented with antibiotics (100 units / mL penicillin and 100 µg / mL streptomycin) and 2 mM glutamine. Infected cell cultures were incubated at 37C until they showed widespread (100%) cytopathic effects (CPE). Virus was harvested, aliquoted and used for the dsRNA extraction.
Extraction of Viral RNA
Viral RNA was purified from BTV–9 infected BHK–21 cells (which were pelleted down by centrifugation at 3000 rpm for 15 minutes) using TRIZOL reagent (Invitrogen, India) as per manufacturer’s instructions. Briefly, the infected cell pellet was lysed in 1 ml of commercially available TRIZOL® reagent (Invitrogen), 0.2 volume of chloroform was added, mixed by vortexing and the mixture incubated on ice for 10 min. The supernatant, containing total RNA, was separated from cellular debris and DNA by centrifuging at 10,000 x g for 10 min at 4°C. An equal volume of isopropanol containing 1/10th volume of 3M Sodium acetate was added to the supernatant, then mixed and the RNA was allowed to precipitate for a minimum of 2 hours at –20°C. The RNA was pelleted by centrifugation at 10,000 x g for 10 min, washed with 70% ethanol, air dried and suspended in nuclease free water (NFW). The RNA was either used immediately or stored at –20°C.
Design of Primer Pairs for Amplification of Genome Segment 2 of BTV–9
Thirty one available sequences of segment 2 of eastern topotype of BTV–9 were obtained from Genbank with Bluetongue virus 9 Indian isolate BBF strain as reference sequence (Accession No. JF443167). Four primer pairs having few degenerate bases to cover the whole range of eastern group of BTV–9 viruses were designed from aligned sequences of segment 2 of BTV–9 using ‘Primer BLAST software’ (http://www.ncbi.nlm.nih.gov/tools/primer–blast/), which are given in Table 1. These primer pairs were excepted to generate four overlapping fragments (A, B, C, D) of 1126 bp, 1105 bp, 867 bp and 856 bp respectively for amplification of full length genome segment 2 of BTV–9 (2931 bp).
One Step Reverse Transcription–Polymerase Chain Reaction (RT–PCR)
Reverse transcription and amplification of genome segment 2 of BTV–9 in four overlapping fragments was performed using Super Script III one step RT–PCR kit from Invitrogen with respective primer pairs. Each PCR reaction was performed in a 25 μL volume in thin walled PCR tubes. Initially, 2 μL of RNA template and 0.5 μL of each primer for a particular set were mixed thoroughly, heat denatured at 95°C for 5 min and snap chilled on ice. Then 12.5 μL of 2x master mix, 0.5 μL of one step RT enzyme was added to the reaction mix. The final volume of the reaction was adjusted to 25 μL by adding 9.0 μL of nuclease free water (NFW). PCR tubes were then spinned and transferred to thermal cycler (Applied Biosystems). RT–PCR reaction was performed using the following reaction conditions: Reverse transcription at 55°C / 30 min, initial denaturation at 95°C / 2 min, followed by 40 cycles of denaturation at 94°C / 15 sec, annealing at 50°C / 30 sec, extension at 68°C / 90 sec with a final extension step of 68°C / 5min. Amplified products were analysed electrophoretically using 1% agarose.
Purification of Amplified Products from Gel
Four segment 2 amplicons were purified using gel extraction kit (Qiagen, India) as per manufacturer’s instructions.
Sequencing Of Segment 2 of BTV–9
The purified PCR amplicons were sequenced directly using ABI PRISMTM 3130 XL Genetic Analyzer (Applied Biosystems) using PCR primers as sequencing primers along with some additional sequencing primers wherever required. Sequence data generated was assembled to make a contiguous sequence of full length segment 2 of BTV–9 using DNA star version 5 of the Lasergene suit. The full length sequence of genome segment 2 of BTV–9 was submitted to Genbank under accession number KJ679573.
RESULTS AND DISCUSSIONS
Bluetongue is regarded as a globally emerging disease and is endemic in India. Outbreaks of BT can be economically devastating to livestock production and the presence of BTV in a country can adversely impact the trade and movement of livestock. Early diagnosis, rapid serotyping and characterization of disease agent (BTV) are essential steps in the identification of origin of the virus and for controlling the disease, which is the first and foremost need to maintain good health of animal flock. Sequencing of genome segment 2 from samples can reliably type the virus strain that is circulating in field. Amplification and sequencing of segment 2 of ~ 3 Kb size is generally difficult from clinical samples as sometimes the clinical material is not available in sufficient amount. Hence, the amplification and sequencing of four overlapping amplicons of genome segment 2 and typing of an isolate of BTV using ABI sequencing platform can be of great use.
All four overlapping fragments (A, B, C, D) of BTV–9 genome segment 2 were successfully amplified giving a single sharp thick band of expected sizes (Table 1) under the optimized conditions of RT–PCR. Figure 1 shows amplification of full length genome segment 2 of BTV–9 of 2931 bp in four overlapping fragments: fragment A – 1126 bp, fragment B – 1105 bp, fragment C – 867 bp, and fragment D – 856 bp respectively on 1% agarose gel electrophoresis.
Figure 1: PCR amplified product of segment 2 of BTV–9 with respective primer pairs (Table 1); Lane L: 1 Kb plus ladder; Lane 1: Pd A – 1126 bp; Lane 2: Pd B – 1105 bp; Lane 3: Pd C – 867 bp; Lane 4: Pd D – 856 bp
Purified products from these bands were cycle sequenced using respective PCR primers as sequencing primers as well as using additional internal primers for sequencing. The sequences obtained from four amplicons were assembled into a single contiguous sequence of 2931 bp representing full length genome segment 2 (shown as line diagram in Figure 2). The sequence obtained was used for Blast analysis to find out the BTV type (http://blast.ncbi.nlm.nih.gov/Blast.cgi). The virus used in this study was typed as BTV–9e showing 99% nucleotide sequence identity with other Indian strains of BTV–9 of eastern lineage. Detailed phylogenetic analysis of the obtained sequence is not presented here as this is not the purpose of this paper.
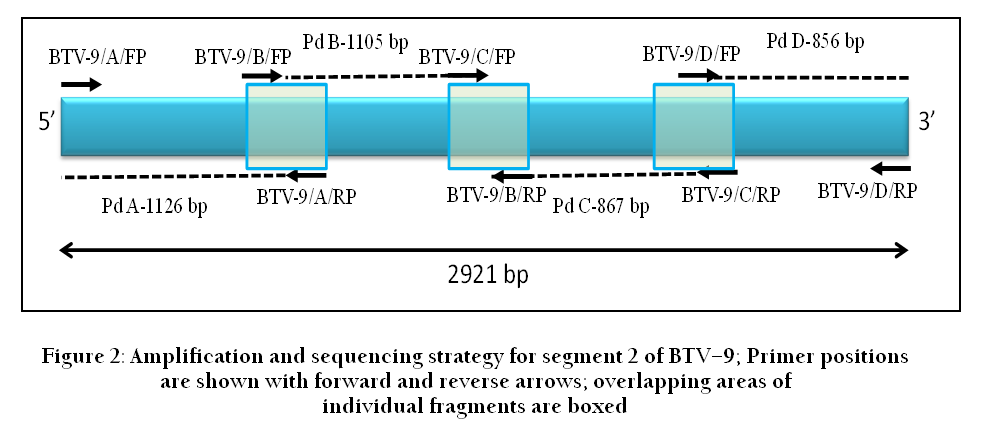
Figure 2: Amplification and sequencing strategy for segment 2 of BTV–9; Primer positions are shown with forward and reverse arrows; overlapping areas of individual fragments are boxed
Molecular typing is replacing serotyping for BTV serotype identification because of limited availability of reference sera and also cross reactions between serotypes. However, it is clear that sequence–based diagnostic assays need to be periodically re–evaluated because there can be random variations in sequences of primer foot prints. VP2 sequences of different strains belonging to the same serotype but separated by space or time may differ significantly. The methodology described here can also be applied to other genome segments of any BTV isolate and even other dsRNA virus genome segments.
CONCLUSION
In this study we have developed an alternative amplification and sequencing strategy for full length genome segment 2 of BTV–9. This application is particularly useful when the initial concentration of RNA in a sample is too low to apply FLAC and/or cloning for obtaining complete gene sequences. Additionally, this methodology can also be applied to other genome segments of any BTV isolate and even other dsRNA viruses. This technique is particularly useful to obtain sequence data from clinical samples where concentration of RNA is very low.
ACKNOWLEDGEMENTS
This work was funded by Rastriya Krishi Vikas Yojna scheme no. 4011/C (g) ABT–4–0A. The authors are thankful to Director of Research and Dr. Trilok Nanda for providing all required facilities. We are also thankful to members of AMRG at the Pirbright Institute, UK for their all support. The assistance provided by non–teaching staff particularly Mr. Chandan Singh is also highly appreciated.
AUTHOR’S CONTRIBUTIONS
SM, NSM: conception and design of experiments, analysis of data, drafting the manuscript, provided reagents and facilities. Sunayna, MS, AG, KB, AK: acquisition of data. All authors have read and approved the manuscript.
CONFLICT OF INTERSEST
We declare no conflict of interest.
REFERENCES
Belhouchet M, Mohd Jaafar F, Firth AE, Grimes JM, Mertens PPC, Attoui H (2011). Detection of a Fourth Orbivirus Non–Structural Protein. PLoS ONE 6(10): e25697. doi:10.1371/journal.pone.0025697.
http://dx.doi.org/10.1371/journal.pone.0025697
Maan NS, Maan S, Nomikou K, Prasad G, Singh KP, Belaganahalli MN, Mertens PP (2012). Full genome sequence of bluetongue virus serotype 1 from India. J Virol 86:4717–4718.
http://dx.doi.org/10.1128/JVI.00536-12
http://dx.doi.org/10.1128/JVI.00420-12
http://dx.doi.org/10.1128/JVI.00731-12
http://dx.doi.org/10.1128/JVI.00671-12
http://dx.doi.org/10.1128/JVI.01672-12
http://dx.doi.org/10.1128/JVI.00188-12
http://dx.doi.org/10.1128/JVI.00596-12
Maan S, Maan NS, Nomikou K, Batten C, Antony F, Belaganahalli MN, Samy AM, Reda AA, Al–Rashid SA, El Batel M, Oura CA, Mertens PP (2011). Novel bluetongue virus serotype from Kuwait. Emerg Infect Dis 17:886–889.
http://dx.doi.org/10.3201/eid1705.101742
PMid:21529403 PMCid:PMC3321788
Maan S, Rao S, Maan NS, Anthony SJ, Attoui H, Samuel AR, Mertens PP (2007). Rapid cDNA synthesis and sequencing techniques for the genetic study of bluetongue and other dsRNA viruses. J Virol Methods 143:132–139.
http://dx.doi.org/10.1016/j.jviromet.2007.02.016
PMid:17433453
Mellor PS, Carpenter S, White DM (2009). Bluetongue virus in the insect host. In: Mellor PS, Baylis M, Mertens PPC, editors. Bluetongue. London Elsevier/Academic Press p295–320.
http://dx.doi.org/10.1016/B978-012369368-6.50018-6
Prasad G, Sreenivasulu D, Singh KP, Mertens PPC, Maan S (2009). Bluetongue in the Indian subcontinent. In: Mellor PS, Baylis M, Mertens PPC, editors. Bluetongue, Ist ed. London Elsevier/Academic Press p167–196.
Rao PP, Reddy YN, Ganesh K, Nair SG, Niranjan V, Hegde NR (2013a). Deep sequencing as a method of typing bluetongue virus isolates. J Virol Methods 193:314–319.
http://dx.doi.org/10.1016/j.jviromet.2013.06.033
PMid:23831448
Rao PP, Reddy YV, Hegde NR (2013b). Isolation and Complete Genome Sequencing of Bluetongue Virus Serotype 12 from India. Transbound Emerg Dis.
Verwoerd DW, Huismans H (1972). Studies on the in vitro and the in vivo transcription of the bluetongue virus genome. Onderstepoort J Vet Res 39:185–191
PMid:4352125


