
Advances in Animal and Veterinary Sciences


Short Communication
Advances in Animal and Veterinary Sciences. 1 (4S): 54 – 58Special Issue-4 (Progress in Research on Viruses and Viral Diseases)
Molecular Characterization of Fowl Pox Virus Isolates from Backyard Poultry
Bithi Roy1, Siddhartha Narayan Joardar1*, Indranil Samanta1, Pradip Kumar Das2, Arkendu Halder1, Sudip Nandi1
- Department of Veterinary Microbiology, Faculty of Veterinary and Animal Sciences, West Bengal University of Animal and Fishery Sciences; P.O.–Belgachia, Kolkata–700037, India
- Department of Veterinary Physiology, Faculty of Veterinary and Animal Sciences, West Bengal University of Animal and Fishery Sciences; P.O.–Belgachia, Kolkata–700037, India
*Corresponding author:joardar69@gmail.com
ARTICLE CITATION:
Roy B, Joardar SN, Samanta I, Das PK, Halder A and Nandi S (2013). Molecular characterization of fowl pox virus isolates from backyard poultry. Adv. Anim. Vet. Sci. 1 (4S): 54 – 58.
Received: 2013–11–04, Revised: 2013–12–25, Accepted: 2013–12–28
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/174/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
With the aim of characterizing the fowl pox field isolates with molecular techniques, scab materials (of comb, wattle and skin of the face) were collected from ten (10) clinically affected backyard chickens from two different agro–climatic zones of West Bengal state, India, viz. Red latterite (Jhargram, District– Paschim Medinipur) and Coastal saline (Kakdwip, District– South 24 Parganas).Viral DNA was extracted from the collected scabs and was subjected to fowl pox virus (FPV) specific Polymerase chain reaction (PCR) for amplification of 4b core protein gene for confirmation of the presence of virus in clinically positive samples. In PCR, an expected amplicon of 578 bp was obtained in all the ten clinically positive samples and in vaccine strain as positive control. Restriction endonuclease analysis (REA) of PCR products with EcoR V and Mse I showed similar restriction patterns among the FPV isolates. Nucleotide sequence analysis of the amplified fragments revealed 99–100% nucleotide similarity with pox virus of other avian species. The present study revealed the molecular characteristics of FPV, circulating in backyard poultry in eastern Indian state West Bengal that was not reported earlier.
Fowl pox is a slow–spreading viral disease of domestic poultry, caused by fowl pox virus (FPV) which is the prototypic species of the genus Avipoxvirus (APV) belonging to the Chordopoxvirinae subfamily of the Poxviridae family (Murphy et al., 1995). The disease is characterized by the presence of discrete, wart–like, proliferative lesions on the skin (cutaneous form) and/or development of moist, necrotic lesions on the mucous membranes of the mouth and upper respiratory tract (diphtheritic form). Mortality in fowl pox is mainly due to this diphtheritic form which causes death by asphyxiation (Tripathy et al., 1978; Sawale et al., 2012). The FPV spreads only horizontally through aerosol and poultry house dust generated by feathers and dried scabs, although occasionally, insects were also implicated in the environmental spread of the disease (Jarmin et al., 2006). FPV can withstand extreme environmental conditions, particularly desiccation, sometimes surviving on perches and in dried scabs for months and years (Tripathy, 1993).
The conventional laboratory diagnosis of FPV is carried out by histopathological examination, electron microscopy, virus isolation on chorioallantoic membrane (CAM) of embryonated chicken eggs or cell cultures, and serologic methods (Siddique et al., 1997; Tripathy and Reed, 1998; Tadese and Reed, 2003; Wang et al., 2006; Islam et al., 2008). For rapid and routine diagnosis, polymerase chain reaction (PCR) has proved to be the most sensitive techniques (Lee and Lee, 1997; Tadese and Reed, 2003; Weli et al., 2004; Luschow et al., 2004; Singh et al., 2007; Jarvi et al., 2008; Biswas et al., 2011; Balachandran et al., 2012). PCR in combination with restriction endonuclease enzyme analysis (REA) followed by sequence analysis of the amplified fragments are used for detection, differentiation and molecular characterization of fowl pox virus isolates (Coupar et al., 1990; Tadese et al., 2003; Luschow et al.,2004).
FPV is currently considered as an emerging pathogen with world–wide distribution. Infection with the fowl pox virus will cause the chickens to have poor growth, poor feed conversion and a precipitous fall in egg production, which is responsible for substantial economic losses to backyard poultry farming. A number of fowl pox outbreaks have been reported from different parts of India including West Bengal state, India. Previous workers have reported the presence of persistent FPV infection in vaccinated and unvaccinated poultry flocks (Dhanesar et al., 1980; Garg et al., 1984; Joshi et al., 1995; Mishra and Mallik, 1996; Dana et al., 2000; Biswas et al., 2011). However, there are very few reports on molecular characteristics of FPV circulating in backyard poultry, though needs attention for better understanding regarding its epidemiology. This paper describes first time the molecular characteristics of FPV in chickens reared in backyard system of management in two different agro climatic zones of West Bengal state.
In the present study, field samples of fowl pox virus (FPV) were collected from infected Rhode Island Red (RIR) chickens, reared in backyard system, from two different agro climatic zones of West Bengal, viz. Red latterite (Jhargram area, District–Paschim Medinipur) and Coastal saline (Kakdwip area, District– South 24 parganas) during March, 2013. Birds were of different age groups (both growers and adults) and they were not vaccinated against fowl pox. Scab materials were collected aseptically from comb, wattle and skin of the face of the birds (n=10) and kept in screw capped vials containing 50% buffered glycerine and stored at –20oC for laboratory examination.
The collected materials were triturated in phosphate buffered saline (pH 7.2) containing antibiotics (penicillin @10,000 IU/ml, streptomycin @500 µg/ml and gentamicin @250 µg/ml) by using sterile mortars and pestles with sand. Following sonication [at 50% amplitude (Hz), 0.5 cycle for 60 sec with an interval of 2 min cooling on ice between each cycle, for 3 times], it was centrifuged at 1300g at 4oC for 30 min and the supernatant was collected and passed through a 0.45 µm filter. The filtrate was checked for sterility and stored at –20oC. This filtrate (100µl) was used for inoculation of specific pathogen free (SPF) embryonated chicken eggs of 10 days old via the route of chorioallantoic membrane (CAM). The eggs were incubated at 37oC and observed for 7 days.
Viral DNA was extracted from 25 mg of the scab materials (field samples), infected (experimentally inoculated) CAM materials of all the collected clinical samples (n=10) and from 25 mg of the live attenuated FPV vaccine (Freeze dried), procured from a commercial firm (Bio Med Private Limited, India) following manufacturer’s guidelines of DNA extraction kit (Genetix, Asia). The DNA concentration was estimated by using Bio–photometer (Eppendrof, Germany).
FPV specific PCR was performed using a primer pair described by Lee and Lee (1997). The sequence of the primers were as follows– (F: 5’–CAGCAGGTGCTAAACAACAA–3’) and (R: 5’–CGGTAGCTTAACGCCGAATA –3’). The primers were based on FPV 4b gene sequence (Binns et al., 1989). The reaction was performed according to Biswas et al., 2011, with slight modification in 50µl volume containing 10mM Tris–HCl (pH 8.8), 50 mM KCl, 1.5 mM MgCl2, 200µM each dNTP, 1.5 U of Taq DNA polymerase, 8µl DNA, 1 µl each of forward (F) and reverse (R) primer (10 Þmol/μl) and up to 50µl of nuclease free water (NFW). PCR was done in a thermal cycler (Mastercycler, Eppendorf, Germany) under the following condition for 30 cycles: initial denaturation at 94oC for 5 min, subsequent denaturation at 94oC for 1 min, annealing at 60oC for 1 min and extension at 72oC for 1 min. The final extension was done at 72oC for 2min. The size of the amplified DNA fragment using these two primers was expected to be 578 base pair (bp) in length. The amplified products of PCR (8µl) were separated by agarose gel electrophoresis using 1.2% agarose in 0.5 x TBE buffer along with 0.5 μl/ml ethidium bromide at 100 volt for 1 hr with 100bp DNA Ladder. The amplified DNA fragments were analyzed using gel documentation system (UVP, UK).
For molecular characterization of FPV, PCR amplified products of 4b gene (578 bp) of both the representative isolates, one each from Jhargram (F1) and Kakdwip (F7) were subjected to restriction endonuclease analysis (REA) using the enzymes Bam HI, EcoR I, EcoR V, Pst I, Hind III and MseI. The PCR products generated from 4b gene of FPV were purified using PCR purification kit (Fermentas). Purified amplicons were subjected to restriction endonuclease enzyme digestion as per the manufacturer’s protocol, employing specific assay condition and the buffers supplied. For digestion, 0.5 to 1µg of purified PCR product (checked on the gel) was taken in a 10µl reaction mix in a 0.5 ml eppendorf tube, containing 1 µl (10U/µl) of restriction enzyme and 1 µl of 10 X compatible RE buffer, the final volume being made up with nuclease free water (NFW). The reaction was performed at 37oC overnight in water–bath and stopped by storing at –20oC until it was subjected to agarose gel electrophoresis. The resulting fragments were analyzed by 2% agarose gel electrophoresis having ethidium bromide, in 0.5 x TBE buffer at 100V for 1 hour. The gels were examined on UV transilluminator and photographed by Gel Documentation system. The size of various fragments of digested viral DNA was determined using GeneTools Software (version 2.0) and expressed in base pairs (bp).
DNA sequencing was carried out for two isolates of Jhargram(F1 ) and Kakdwip (F7 )by a commercial DNA sequencing service (Genomics Division, Xcelris Labs Ltd, Ahmedabad, India). The nucleotide sequences generated in the present study has been deposited in the Genbank database, NCBI. The sequences were edited, assembled and analyzed with the software GeneFisher 2. Sequences were compared with the published FPV nucleotide sequences with the help of NCBI nucleotide blast.
In Jhargram, six birds (n=6) and in Kakdwip, four birds (n=4) were affected in the flock of 18 and 22 birds, respectively. Small nodules, brownish and blackish in colour (Pock lesions) were observed on the comb, wattle, around the eyelids, skin of the face (non–hairy part) and legs of the diseased birds (Figure 1). Eyelids were swollen and there were accumulation of cheesy exudative deposits under them. In Jhargram, two birds showed pin–head sized light yellowish lesions on the mucosal surface of tongue and soft palate along with the cutaneous lesions. The birds appeared sick with roughened feather and stunted growth. They were unwilling to take food and tend to stand in one place lowering down the head. There was drop in egg production in layer birds.

Figure 1: Chicken showing pock lesion on comb, wattle, around the eyelids and non hairy part of the face
The total amount of viral DNA extracted from each scab material (n=10) as estimated from A260 nm value was between 24.5µg and 56.4µg per ml. The ratio of OD260/280 for isolated DNAs was between 1.70 and 2.00, which indicated the purity of DNA samples. To confirm the presence of FPV in scab materials, FPV specific PCR was performed and an expected size amplicon of 578 bp was obtained (Figure 2) in all the samples (F1– F10), collected on the basis of clinical lesion. PCR amplification of FPV vaccine using the same primers of 4b core protein gene also revealed an amplicon of 578 bp which acted as positive control in PCR analysis.

Figure 2: PCR analysis of FPV 4b core protein gene of FPV field isolates from scab materials showing amplified 578 bp fragments
The CAM of inoculated eggs revealed prominent thickening of the membrane after 7th day post inoculation (PI) in all the samples. FPV specific PCR of all the infected CAM materials also generated expected amplicons of 578 bp that confirmed the presence of FPV (Figure 3a and 3b).
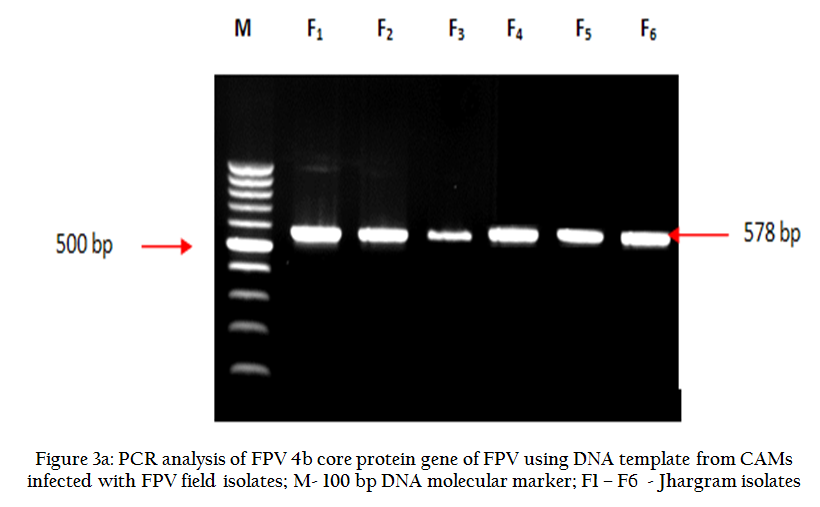
Figure 3(a): PCR analysis of FPV 4b core protein gene of FPV using DNA template from CAMs infected with FPV field isolates
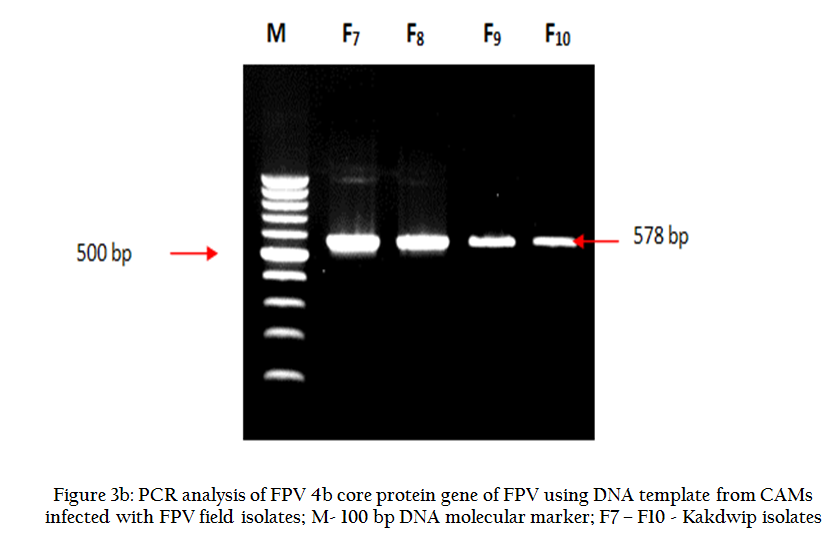
Figure 3(b): PCR analysis of FPV 4b core protein gene of FPV using DNA template from CAMs infected with FPV field isolates
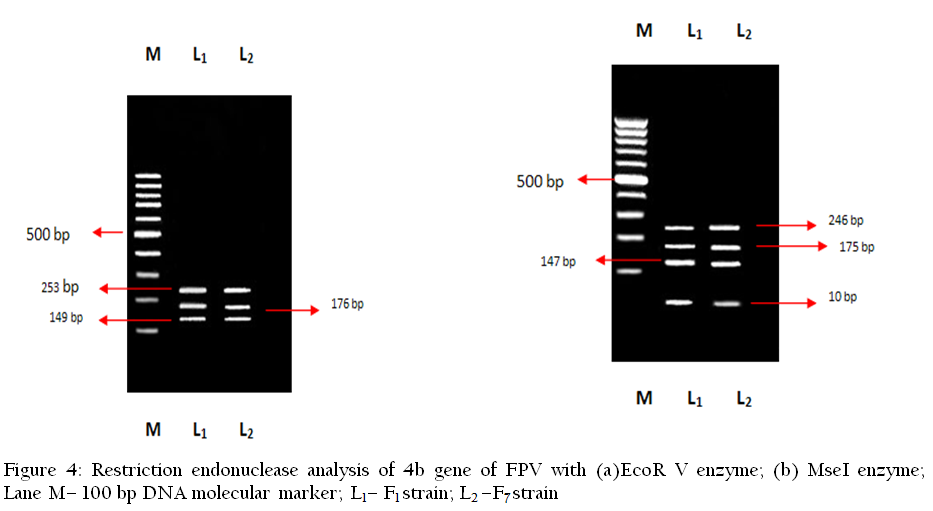
When the 4b gene amplicon of the F1 and F7 isolates of FPV were analyzed by the endonuclease enzymes viz. Bam HI, EcoR I, PstI and Hind III, they remain undigested due to lack of the available restriction site for the enzymes to act. Thus the cleavage sites for these enzymes were found restricted to 4b gene sequences. With the enzyme EcoR V, identical restriction patterns were observed for both the F1 and F7 FPV isolates. When the FPV 4b gene amplicon of both the isolates F1 and F7, were digested with EcoRV enzyme, 3 fragments of 253, 176 and 149 bp were observed, indicating two cutting sites for this enzyme. But when digestion of 4b gene amplicon of F1 and F7 isolates was done with enzyme MseI, both the isoloates (F1 and F7) revealed similar restriction patterns. With MseI, 4 fragments of 246, 175, 147 and 10 bp were observed. Results of RE analysis are shown in Figure 4.
Both the obtained sequences were 578 bp fragment of the 4b gene of the FPV and correspond to the 200750–201327 nucleotides of the FPV, complete genome (GenBank accession number AF198100). The primer sequences used in this study was obtained from FPV (HP444) core protein 4b gene sequence (GenBank accession number M25781). When both the sequences were compared with this 4b gene sequence, 100% homology was observed corresponding to the 458–1035 nucleotides (Table 1). Comparing with few previously published Indian isolates, FPV isolate (Golden Pheasant) 2008/India/01 P4b core protein gene (GenBank accession number HM481402); FPV isolate (Golden Pheasant) 2008/India/02 P4b core protein gene (GenBank accession number HM481403); FPV isolate (Indian Peacock) 2008/India/03 P4b core protein gene (GenBank accession number HM481404); FPV isolate (Indian Peacock) 2008/India/04 P4b core protein gene (GenBank accession number HM481405); FPV isolate (Silver Pheasant) 2008/India/05 P4b core protein gene (GenBank accession number HM481406); and FPV isolate (Sparrow) 2009/India/06 P4b core protein gene (GenBank accession number HM481407), revealed 99% homology. Details of the comparison, with the related nucleotide sequences are shown in (Table–1).The edited alignments for both F1 (Jhargram) and F7 (Kakdwip) were 578 bp in length and these sequences were deposited in GenBank under the accession number KF548037and KF548036.
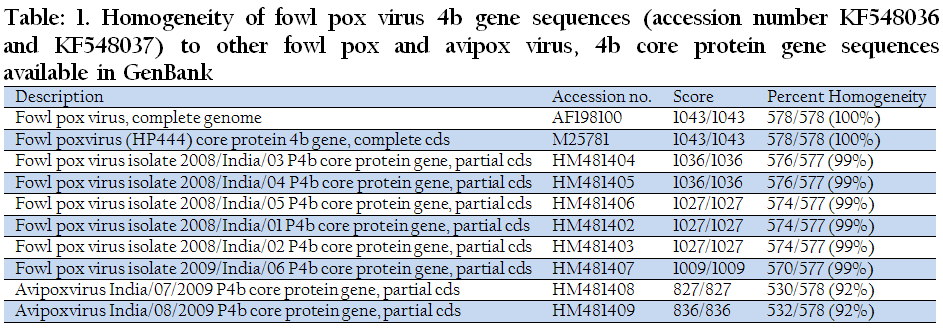
Table 1: Homogeneity of fowl pox virus 4b gene sequences (accession number KF548036 and KF548037) to other fowl pox and avipox virus, 4b core protein gene sequences available in GenBank
Fowl pox is the most common viral disease after Ranikhet disease, encountered by the backyard poultry farmers in West Bengal. It possesses a serious threat among the poor farmers by reducing the productivity to a great extent. This study reports on FPV infection in backyard poultry birds in villages of two different agro climatic zones of West Bengal (Jhargram and Kakdwip). Earlier, FPV was detected based on clinical symptoms in Binpur block of Medinipur district, North 24 Parganas and Jalpaiguri districts of West Bengal (Dana et al., 2000; Saha, 2003; Das et al., 2013). Molecular detection of FPV along with reticuloendotheliosis virus was reported by Biswas et al. (2011) in a flock of backyard chickens in old alluvial zone (Burdwan district) of West Bengal. The present study confirms the presence of Fowl pox virus by PCR in backyard poultry in two different agro–climatic zones of West Bengal. From the present finding, it can be said that routine diagnosis of FPV by PCR is one of the most rapid, reliable and efficient molecular techniques and that needs to be applied for knowing the magnitude of viral infection in backyard poultry. Poor farmers rearing backyard poultry seldom practice vaccination against fowl pox. Earlier, in the year 2011 and 2012, there were history of fowl pox outbreak in the same and/or nearby flocks causing severe production loss. In 2012, mortality was reported in the same flock of Jhargram (personal communication). The present study also indicated the need to generate awareness among the farmers to follow vaccination schedule of fowl pox in backyard flocks to combat recurring economic losses associated with the disease.
Identical restriction pattern was observed for both the isolates (F1 and F7) when restriction enzyme analysis by EcoR V was performed. In both cases, 3 fragments of 253, 176 and 149 bp were observed, indicating two cutting sites for this enzyme. Similarly, identical RE pattern was seen in both the cases when MseI was used. Here, 4 fragments of 246, 175, 147and 10 bp were observed. This shows that the isolates are quite similar at least based on the molecular characteristics. That means similar strain of virus was circulating in two different agro–climatic zones of West Bengal. Nucleotide sequence analysis of the isolates reveals 99% similarity with the previously isolated strains of India from golden pheasants, Indian peacock, silver pheasant and sparrow. This shows that the present isolated strain is widely prevalent in different avian species in different geo–climatic regions. The present finding corroborates the previous report of Luschow et al. (2004) where no differentiation was found among FPV and poxvirus isolated from turkey and sparrow upon nucleotide sequencing and phylogenetic analysis.
PCR in combination with REA and sequence analysis is an effective system to characterize FPV, circulating in backyard poultry and as such more elaborate studies in future encompassing these parameters are suggested for better understanding the epidemiology of FPV in backyard system of management.
The present study revealed the molecular characteristics of Fowl pox virus circulating in backyard poultry of eastern Indian state West Bengal.
ACKNOWLEDGEMENTS
The work was supported partially by Rastriya Krishi Vikas Yojona (RKVY) scheme, Govt. of India. Authors wish to thank The Vice Chancellor, West Bengal University of Animal & Fishery Sciences, Belgachia, Kolkata for providing necessary research facilities.
REFERENCES
Balachandran C, Pazhanivel N, Prabhakar TG, Murugadas V and Prabakar P (2012). Avipox virus infection in rosella parakeet (Platycercus sp.). J.Adv.Vet. Res.2: 184–187
Binns MM, Boursnell MEG, Tomley FM and Campbell J (1989). Analysis of the fowlpoxvirus gene encoding the 4b core polypeptide and demonstration that it possesses efficient promoter sequences. Virol. 170: 288–291.
http://dx.doi.org/10.1016/0042-6822(89)90380-2
Biswas SK, Jana C, Chand K, Rehman W and Mondal B (2011). Detection of fowlpox virus integrated with reticuloendotheliosis virus sequences from an outbreak in backyard chickens in India. Vet. Ital. 47(2):147–153.
PMid:21706467
Coupar BE, Teo T, Boyle DB (1990). Restriction endonuclease mapping of the fowlpox virus genome. Virol.179: 159–167.
http://dx.doi.org/10.1016/0042-6822(90)90285-Y
Dana SS, Rathore BS and Kaul PN (2000). Morbidity and mortality pattern in desi chicken reared by the santal tribe of west Bengal. Indian J. Anim. Res. 34(1): 49–51.
Das P, Joardar SN, Samanta I, Das PK, Jana C, Isore DP and Sadhukhan TK (2013). Isolation and molecular confirmation of Avipoxvirus in backyard chicken in West Bengal. Indian J. Comp. Microbiol. Immunol. Infect. Dis. (in press).
Dhanesar NS, Sharma SN and Malik BS (1980). Fowlpox outbreaks in vaccinated flocks and isolation of virus strains. Indian Vet. Med. J. 4: 144–148.
Garg SK, Joshi HC and Shukla HC (1984). Vaccination failure in fowlpox. Indian J. Vet. Med. 4: 60–61.
Islam MR, Khan MSR, Islam MA, Kayesh MEH, Karim MR, Gani MO and Kabir A (2008). Comparative efficacy of imported fowl pox virus vaccine with locally produced one in backyard chicks. Bangl. J. Vet. Med.6 (1):23–26.
http://dx.doi.org/10.3329/bjvm.v6i1.1334
Jarmin S, Manvell R, Gough RE, Laidlaw SM, Skinner MA (2006). Avipoxvirus phylogenetics: identification of a PCR length polymorphism that discriminates between the two major clades. J. Gen. Virol. 87: 2191–2201.
http://dx.doi.org/10.1099/vir.0.81738-0
PMid:16847115
Jarvi SI, Triglia D, Giannoulis A, Farias M, Bianchi K, Atkinson CT (2008). Diversity, origins and virulence of Avipoxviruses in Hawaiian forest birds. Conserv.Genet. 9 (2): 339–348.
http://dx.doi.org/10.1007/s10592-007-9346-7
Joshi RK, Shakya S and Gupta N (1995). Isolation of fowl pox virus from fowl pox vaccinated layer flock. Indian J. Comp. Microbiol. Immunol. Infect. Dis. 16: 139–140.
Lee LH and Lee KH (1997). Application of the polymerase chain reaction for the diagnosis of fowlpox virus infection. J. Virol. Methods 63: 113–119.
http://dx.doi.org/10.1016/S0166-0934(96)02119-2
Luschow D, Hoffmann T and Hafez HM (2004). Differentiation of avian poxvirus strains on the basis of nucleotide sequences of 4b gene fragment. Avian Dis.48: 453–62.
http://dx.doi.org/10.1637/7111
PMid:15529967
Mishra SS and Mallik BB (1996). Comparative immunological and genomic characterization of fowlpox virus isolates. Indian J. Exp. Biol. 34: 11–17.
PMid:8698401
Murphy FA, Fauquet CM, Bishop DHL, Ghabrial SA, Jarvis AW, Martelli GP, Mayo MA, Summers MD (1995). Avipoxvirus. In: Virus Taxonomy, Classification and Nomenclature of Viruses, Springer, New York, NY, pp. 85.
Saha D (2003). M.V.Sc Thesis, Indian Veterinary Research Institute. Izatnagar, Bareilly, U.P.
Sawale GK, Roshini S, Bulbule NV, Chawak MM and Kinge GS (2012). Pathology of fowl pox in chickens. Indian J. Vet. Pathol. 36(1): 110–111.
Siddique B, Rahman MB, Amin MM and Rahman MM (1997).Antibody titres in chicks following pigeon pox virus inoculation. Bangl. Vet. J. 14 (1–2): 12–14.
Singh A, Dash BB, Kataria JM and Dhama K (2007). Application of polymerase chain reaction and restriction endonuclease analysis for the detection and differentiation of turkey–pox and fowl–pox virus infections. Indian J. Anim. Sci. 77: 521–525.
Tadese T, Potter EA and Reed WM (2003).Development of a mixed antigen agar gel enzyme assay (AGEA) for the detection of antibodies to pox virus in chicken and turkey sera. J. Vet. Med. Sci. 65: 255–58.
http://dx.doi.org/10.1292/jvms.65.255
PMid:12655123
Tadese Tand Reed WM (2003). Detection of specific reticuloendotheliosis virus sequence and protein from REV–integrated fowlpox virus strains. J.Virol. Methods 110: 99–104.
http://dx.doi.org/10.1016/S0166-0934(03)00106-X
Tripathy DN (1993). Avipoxviruses. In: Virus Infections of Vertebrates − Virus Infections of Birds. McFerran JB and McNulty MS (eds.). Elsevier Science Publishers, Amsterdam, Netherlands. pp. 5–15.
Tripathy DN and Hanson LE (1978). Pathogenesis of fowl pox in laying hens. Avian Dis. 22: 259–265.
http://dx.doi.org/10.2307/1589537
PMid:209778
Tripathy DN and Reed WM (1998). Pox. In: A Laboratory Manual for the Isolation and Identification of Avian Pathogens. Swayne DE, Glisson JR, Jackwood MW, Pearson JE and Reed WM (eds.). American Association of Avian Pathologists, University of Pennsylvania, New Bolton Center, Kennett Square, PA, USA. pp. 137–140.
Wang J, Meers J, Spradbrow PB and Robinson WF (2006).Evaluation of immune effects of fowl pox vaccine strains and field isolates.Vet.Microbiol.116: 106–119.
http://dx.doi.org/10.1016/j.vetmic.2006.03.012
PMid:16650660
Weli SC, Okeke MI, Tryland M, Nilssen O and Traavik T (2004). Characterization of Avipoxviruses from wild birds in Norway. Canadian J. Vet.Res.68:140–145.
PMid:15188959 PMCid:PMC1142158

