
Advances in Animal and Veterinary Sciences


Review Article
Advances in Animal and Veterinary Sciences. 1 (2S): 7 – 14Special Issue – 2 (Clinical Veterinary Practice–Trends)
An Overview of Mechanisms and Emergence of Antimicrobials Drug Resistance
Sujeet Kumar*, Bhoj Raj Singh
-
Indian Veterinary Research Institute, Izatnagar, Bareilly, India
*Corresponding author:sujeetmicrobiol@gmail.com
ARTICLE CITATION:
Kumar S and Singh BR (2013). An overview of mechanisms and emergence of antimicrobials drug resistance. Adv. Anim. Vet. Sci. 1 (2S): 7 – 14.
Received: 2013–09–16, Revised: 2013–10–24, Accepted: 2013–10–24
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/121/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Emergence of antimicrobial drug resistance in bacteria is widely explored but still ill understood. Antibiotic resistance is a complex phenomenon due to multiple ways of its acquisition, mechanism of action and spread. Antibiotic resistance may be innate, present in microbes due to their inherent inbuilt metabolic and structural components. Other type of innate resistance is one which has been detected in strains isolated even before the era of the use of any antibiotics and often specific with bacterium. The most important and problematic is the acquired drug resistance. It spread rapidly with the use of antimicrobials in one or other realm of ecosystem. Inactivation of antimicrobials occurs by some common mechanism such as drug inactivation/degradation by bacterial enzymes or alteration in the bacterial targets or expulsion of drug out of the bacterial cells (efflux) or by preventing the entry of drug into bacterium. Here some important aspects of mechanism of antimicrobial drug resistance and its emergence are discussed.
INTRODUCTION
The field of medical and veterinary sciences was revolutionized by the discovery of antibacterial drug by the middle of the 20th century. It resulted to dramatic improvement in human and animal health and productivity. The diseases like tuberculosis, bubonic plague, cholera which were the scourge throughout the human civilization came under complete control. Soon, scientists and planner entered in euphoria that bacterial diseases will be just a history in very short time (Singh, 2011, 2013). However, euphoria remained short–lived, as bacteria responded against the antibiotics by manifesting different forms of resistance. The situation is rapidly turning now in the favor of bacterial pathogens. The number of new antibacterial agents has started to dwindle. But bacteria are evolving day over the day more and more clever mechanisms of resistance, and posing a serious threats to human and animal health (Levy and Marshall, 2004; Singh, 2013).
Use of Antimicrobials in Animals
In animals antibiotics are used for the following purposes (Schwarz et al., 2001):
Therapeutic uses: To treat infections and applied in therapeutic doses (higher than used for preventive, control and production purposes) and for a short period.
Preventive or control uses: For prevention of a disease, to control the spread of one or other disease in a situation where disease is likely to occur due to environmental conditions and exposure of disease causing organisms.
Production or growth promotion uses: It is to increase daily body weight gain and/ or to enhance feed efficiency by administering doses lower than those required to treat or prevent disease over an extended period of time. Often designated as nonprescription use.
According to recent estimates, 80% of total antibiotics produced in world are used in animals. In USA total human antibiotic use was estimated at 8 billion pounds per annum while in animals it amounts to record 29.9 billion pounds per year (Marshal and Levy, 2011). During 2006, China used 97,000 tons of antibiotics in animal feed out of total 210,000 tons of production. Today China produces almost 400,000 tons of antibiotics annually and most of the production is meant for animal husbandry (Li et al., 2013a). Most of the antibiotics used in human or animals as therapeutic drugs have been used as antibiotic growth promoters (AGPs) in one or other kind of animal farming. Evidences suggest the role of AGPs in emergence of multiple drug resistance (Singh, 2011). In view of this, Sweden first took the initiative to prohibit the use of antimicrobials in animal feed. European Union and USA restricted or banned several antimicrobials for non–prescription use in animals and aquaculture. The commonly used antimicrobials used as AGPs include, amoxicillin, ampicillin (AGPs in aquaculture), ardacin (bovine AGP, a glycopeptides withdrawn from EU in 1997), avilamycin (AGP for broilers withdrawn from EU), avoparcin (AGP, a glycopeptides withdrawn from EU in 1997), bacitracin (AGP for poultry, swine, and beef cattle, withdrawn from EU in 1999 but available in USA), bambermycin (AGP for poultry, swine and cattle, withdrawn from EU in 2006 but available in USA), colistin (in broiler, swine and cattle feed in Japan), efrotomycin (AGP for swine), erythromycin (AGP for poultry, cattle and swine), lasalocid (AGP for cattle, poultry, sheep and rabbits approved in EU and USA), lincomycin (AGP for chicken and swine, approved in USA), monensin (AGP in bovine and poultry, withdrawn from EU for bovine use but authorized as poultry coccidiostat), neomycin (AGP for swine and poultry approved in USA), narasin (poultry feed coccidiostat, AGP for cattle approved in USA), nourseothricin (swine AGP withdrawn from EU), olaquindox (swine AGP, withdrawn due to toxicity in EU and Canada, available in USA), oleandomycin (poultry and swine AGP), ormetoprim (poultry AGP for prevention of fowl cholera and other infections), pristinamycin (AGP for wide use), procaine penicillin (AGP in poultry and swine, withdrawn in USA), roxarsone (AGP for poultry and swine), salinomycin (swine AGP), spiramycin (swine AGP withdrawn from EU in 1999), sulfonamides (aquaculture, swine, chicken AGP), tetracycline (AGP for poultry, swine and cattle withdrawn from EU, authorized in USA), tiamulin (swine AGP), tylosin (swine AGP, withdrawn from EU, available in USA) and virginiamycin (AGP for broilers, withdrawn from EU and available in USA) (Samanidou and Evaggelopoulou, 2008).
Excessive and unnecessary use of antimicrobials, particularly as growth promoters in livestock and poultry may lead to a great risk to human health (WHO, 2013). In 1998, the European Union banned the use of antimicrobials for growth promotion in animals as per WHO recommendations. An increase in international trade of animals and animal products is also a concern in the spread of antibiotic resistance. (Tiwari et al., 2013a; WHO, 2013). To overcome the hurdles of antibiotics resistance various alternative emerging novel therapies are coming into picture such as herbal medication, ethno–veterinary medicines, bacteriophage therapy, cytokine therapy, mycophage therapy, panchgavya therapies etc. which are opening new avenues to fight against these superbugs (Dhama et al., 2013a, 2013b; Tiwari et al., 2013a, 2013b).
Antibiotics and their Spectrum
The term antibiotic was coined by Selman Waksman, a Nobel–laureate soil microbiologist who discovered streptomycin. Antibiotic is defined as a compound produced by a microbe which kill or inhibit the growth of another microbe. Hundreds of antibiotic molecules of natural, semi–synthetic and synthetic nature are used for treating infections. The mechanism of action by these antimicrobial compounds is summarized in Table 1. The different antimicrobials used in veterinary medicine and their mechanism of action has been has been extensively reviewed by Schwarz and Chaslus–Dancla (2001) and Andre and Bryskier (2005).
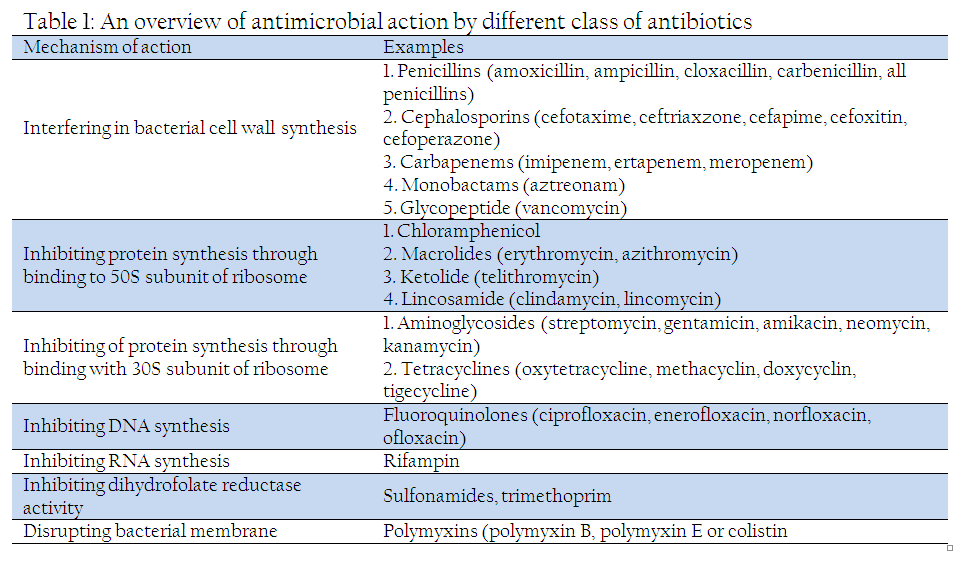
History of Antibiotic Resistance
Sulfonamide was introduced in 1937 as a first effective antimicrobial agent. Since then the development of resistance has plagued their therapeutic use. Penicillin was discovered by renowned microbiologist Alexander Fleming in 1928. But before its introduction as a therapeutic agent the enzyme (bacterial penicillinase/ β–lactamases) which destroy it was identified (Abraham and Chen, 1940). Later methicillin was developed by chemically modifying the penicillin to prevent cleavage by β–lactamases. But just within 3 years of introduction of the methicillin, the drug resistant bacteria were reported (Davies and Davies, 2010). In 1944, streptomycin was introduced for the treatment of tuberculosis. Now tuberculosis treatment is facing the serious challenge due to development of resistance against streptomycin and most of other antimicrobials used in tuberculosis kit. The whole concept of drug resistance started changing by as early as mid–1950. It became evident that antibiotic resistance is genetically transferable. Later, it was observed that antibiotic resistance genes could be disseminated among the bacterial pathogens by conjugation. Many other mechanisms like transformation and transduction were also observed with respect to antibiotics resistance. In the light of many new findings, the area has been the matter of recent review in many leading journals (Alekshun and Levy, 2007; Allen et al., 2010; Davies and Davies, 2010). In brief, the sequence of antibiotics discovered and emergence of resistance is briefed in Table 2.

Superbugs
The term "superbugs” refers to microbes having high level of drug resistance and causing enhanced morbidity and mortality to no response to therapeutic efforts. Super–bug infections limit the therapeutic options for the clinicians; extend the periods of hospitalization and thus the treatment cost. In some cases, super–resistant strains have also acquired increased virulence and enhanced transmissibility. Over the years, continued selective pressure by different antimicrobials led to multidrug resistance (MDR) in human and animal pathogens. Some of the most problematic MDR organisms include methicillin–resistant Staphylococcus aureus (MRSA), extensively drug–resistant (XDR) Mycobacterium tuberculosis, vancomycin–resistant Staphylococcus aureus (VRSA), vancomycin–resistant enterococci (VRE), Klebsiella pneumoniae bearing extended–spectrum β–lactamases (ESBL), Pseudomonas aeruginosa, Escherichia coli and Acinetobacter baumannii (Foster, 2004; Arias and Murray, 2009; Davies and Davies, 2010).
Tuberculosis infects nearly one–third of the world population. Emergence of MDR strain of M. tuberculosis is seriously compromising the TB therapy (Blanchard, 1996). In the last decade, extremely drug–resistant (XDR) strains resistant to four or more front–line antibiotics used against tuberculosis. The XDR strains are now being reported from several parts of the world and posing severe threat to community health. Moreover, the emergence of total drug resistant (TDR) strains is compelling to think for the coming back to pre–antibiotics era (Alanis, 2005; Davies and Davies, 2010).
The Gram–negative pathogens such as Escherichia coli and Klebsiella pneumoniae cause variety of diseases in humans and animals. Antibiotic resistance has been observed in these pathogens against lactam class of antibiotics. These pathogens show a great deal of diversity and produces upto 1000 different types of β–lactamases (Hancock, 1998).
Pseudomonas aeruginosa has emerged as a major nosocomial threat apart from being involved in burn wound infection. It has evolved resistance against β–lactam and aminoglycosides group of antibiotics (Livermore, 2002; Breidenstein et al., 2011). Acinetobacter baumannii is another Gram–negative pathogen associated with nosocomial infection. It got equipped with all different kind of resistance genes due to their remarkably high ability of natural transformation (Poirel and Nordmann, 2006; Arias and Murray, 2009).
Staphylococcus aureus is regarded as one of the most notorious superbugs (Foster, 2004; Lindsay and Holden, 2004). The pathogen resides as nasal commensal in almost 30% of human population. It is now recognized as a major nosocomial infection. Methicillin was the first anti–resistance antibiotic developed against the penicillinases. But within three years of introduction of methicillin, a methicillin–resistant S. aureus (MRSA) emerged. Due to development of MDR strain of S. aureus, the acronym now denotes multidrug–resistant S. aureus.
Clostridium difficile is a Gram–positive, spore forming and toxin–producing anaerobe. It causes severe intestinal infections as a result of extensive use of antibiotics. The depletion of intestinal microflora by antibiotics and subsequent colonization by C. difficile is thought as primary cause of antibiotic associated diarrhea (AAD) or post–antibiotic diarrhea (Abigail et al., 2011).
Antimicrobial Resistance Mechanism
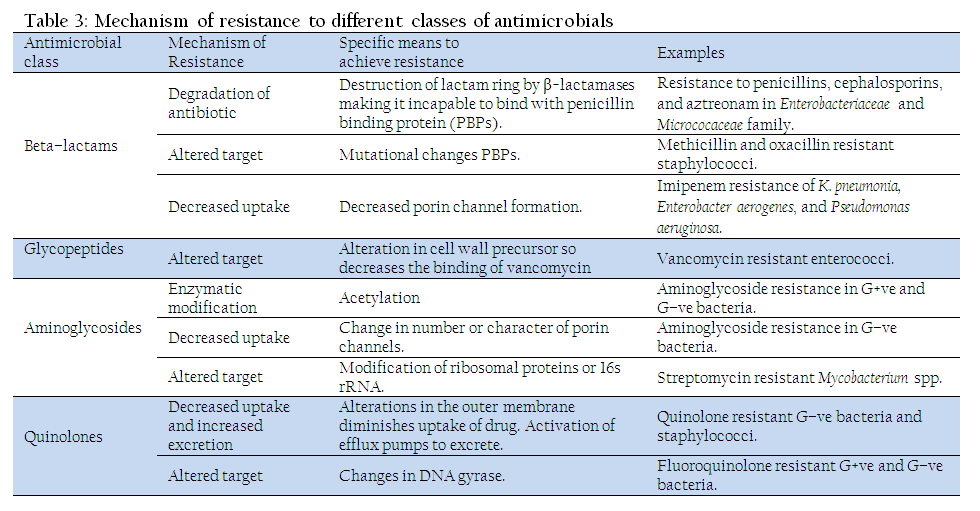
The mechanism of antibiotics resistance has extensively been reviewed (Hawkey, 1998; Alanis, 2005; Higgins, 2007; Alekshun and Levy, 2007; Arias and Murray, 2009; Davies and Davies, 2010). The mechanisms of resistance for different class of antimicrobials have been summarized in Table 3. Several different mechanisms may work together to confer resistance to a single antimicrobial agent. Overall resistance mechanisms adopted by bacteria are classified (Brötz–Oesterhelt and Brunner, 2008) as:
Reduction in entry or access to the target site of antimicrobial drug
Activation of efflux mechanism to expel antimicrobial agents from the bacterial cell
Enzymatic degradation or modification of antimicrobials either inside or outside of the bacterial cell
Modification of antimicrobial drug target(s) within the bacterial cell
Reduction in Entry or Access to the Target Site of Antimicrobial Drug
Antimicrobial compounds require entry and or access into the bacterial cell to interfere with the normal function of bacteria. Porin channels serve as passage for antibiotics to cross the bacterial outer membrane. Few bacteria protect themselves by prohibiting entry of antimicrobials into their cell (Mah and O'Toole, 2001), viz., many Gram–negative bacteria reduce the uptake of aminoglycosides and beta lactam by modifying the cell membrane porin channel numbers, size, and selectivity. This prevents aminoglycosides and beta lactams to reach their intended targets, ribosome and the penicillin–binding proteins (PBPs), respectively. This strategy has been observed in Pseudomonas aeruginosa and Klebsiella spp. against imipenem (a beta–lactam antibiotic), and vancomycin intermediate resistant S. aureus (VISA) strains have thickened cell wall to trap the drug before entry in to the cell (Howden et al., 2010)
Activation of Efflux Mechanism to Expel Antimicrobial Agents from the Bacterial Cell
Efflux mechanism of antibiotic resistance has extensively been reviewed (Poole, 2005; Piddock, 2006; Higgins, 2007). Bacteria possess membrane pumps to move lipophilic or amphipathic molecules across the cell wall. Efflux pump is true variant of these existing pumps. Antibiotic producing bacteria widely use such pump to move antibiotics out of the cells as fast as they are produced. Therefore, it constitutes a protective mechanism to prevent bacteria getting killed by their own chemical weapons. Several pathogenic bacteria possess trans–membrane proteins which act as an efflux pump. This pumps the antibiotic out of the cell as fast as it enters. This results in low intracellular concentrations of antibiotics insufficient to elicit antibacterial effect. The efflux pumps are sometime selectively specific to extrude antibiotics like tetracycline, macrolides, lincosamide, and streptogramins. Many other pumps possess ability to expel multiple drugs which can expel a variety of structurally different antimicrobials (Piddock, 2006). This strategy has been observed in E. coli and other Enterobacteriaceae against tetracycline and chloramphenicol, in staphylococci against macrolides and streptogramins and in S. aureus and Streptococcus pneumoniae against fluoroquinolones (Li et al., 2013a).
Enzymatic Degradation or Modification of Antimicrobials Either Inside or Outside of the Bacterial Cell
Degradation mechanism of antibiotics resistance (Wright, 2005) is the first ever antibiotic resistance mechanism observed shortly after the discovery of penicillin, even before its commercial application (Abraham and Chain, 1940). In the process bacteria destroy the active component of the antimicrobial agent. It is the most common mechanism to inactivate β–lactam group of antibiotic, aminoglycoside and chloramphenicol. A classic example is the hydrolytic degradation of the β–lactam ring in penicillin and cephalosporin by the bacterial β–lactamases. The inactivation product, penicilloic acid, is ineffective in binding to penicillin binding proteins (PBPs), thereby can’t prevent cell wall synthesis. Besides, the strategy has also been used by both Gram positive and Gram negative bacteria to inactivate aminoglycosides and chloramphenicol through acetylation, adenylation and phosphorylation (Davies, 1994; Jana and Deb, 2006).
Modification of Antimicrobial Target Within the Bacterial Cell
Modification in target sites allows some bacteria to avoid recognition by antimicrobial agents (Spratt, 1994). Such strategy has been observed in methicillin resistant S. aureus (MRSA) through change or acquisition of different PBPs, in vancomycin resistant Enterococcus (Walsh and Howe, 2002), in sttreptomycin resistant Mycobacterium (through modification of ribosomal proteins or 16s rRNA), mutations in RNA polymerase lead to rifampicin resistance in M. tuberculosis (Rattan et al., 1998; Rodríguez–Verdugo et al., 2013) and mutations in DNA gyrase lead to resistance for quinolones in many Gram–negative bacteria, S. aureus and Streptococcus pneumonia (Hooper, 2001)
Molecular Mechanism of Antimicrobial Resistance
The molecular mechanism of antibiotics resistance has been extensively reviewed time to time (Blanchard, 1996; Hawkey, 1998; Walsh, 2000; Alanis, 2005; Alekshun and Levy, 2007). The bacterial abilities to adopt various strategies for antibiotic resistance are all genetically encoded. These genetic mechanisms are classified into two types;
- Intrinsic resistance
- Acquired resistance
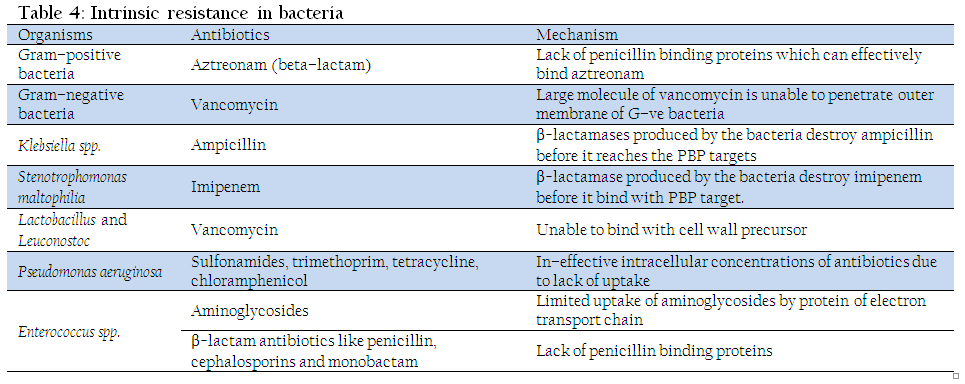
- Intrinsic resistance
It is the innate ability of bacteria to resist antimicrobial effect of particular antibiotic class through its inherent structural or functional characteristics. Such resistance can also be called as "insensitivity” as those microbes have never been susceptible to that particular drug. The classical example is resistance of anaerobes to aminoglycosides and of G–ve bacteria to vancomycin (Fajardo et al., 2008). Different type of intrinsic resistance existing in different class of antibiotics is presented in Table 4. The natural insensitivity can be due to lack of drug targets, inability of drug to enter bacterial cell, expulsion of antimicrobials by chromosomally encoded efflux pump and innate production of antibiotic inactivating enzymes.
- Acquired resistance
It is the ability of bacteria to resist the activity of a particular antimicrobial agent to which it was earlier susceptible. This is mediated by mutation or horizontal gene transfer (transformation, transduction or conjugation) which brings changes in bacterial genome. This brings alteration in the bacterial structural and functional characteristics leading to resistance against a particular antibiotic. The genetic mechanism for acquiring resistance by different antimicrobial class has been summarized in Table 5.

Mutation
Mutation is defined as "spontaneous change in DNA sequence within the gene”. A change within a single nucleotide base pair brings corresponding change in one or more amino acids. This consequently changes the affinity of antimicrobials towards the targeted site.
In bacteria, mutations naturally occur due to errors in DNA polymerase activity, insertions, deletions, and duplications. Due to DNA polymerase error, bacteria have a spontaneous mutation rate of about 0.0033 mutations in each DNA replication cycle. However, the mutation rate varies between the genes.
Mutational alteration in the antibiotic target sites is the reason behind the development of multidrug–resistant (MDR) mycobacterial infections. Streptomycin resistance in Mycobacterium tuberculosis occurs more frequently than in E. coli. Streptomycin works through its binding to 30S sub–unit of ribosome. Therefore, its activity is reduced once mutational alteration happens in ribosomal protein S12 of 16S ribosomal RNA (rRNA). The slow growing bacteria like M. tuberculosis posses only a single copy of rRNA genes in place multiple copies by fast–growing bacteria like E. coli. So, it makes easier for mycobacteria to gain mutational alteration in 16S rRNA or ribosomal protein S12 for resistance against streptomycin. Mycobacterium tuberculosis acquires isoniazid resistance through mutational alteration in katG gene (encoding catalase), inhA gene (target for isoniazid) oxyR gene and aphC gene (Hazbón et al., 2006; Laurenzo and Mousa, 2011). S. aureus
Hydrolytic inactivation of lactam ring by β–lactamases is the most common resistance mechanism in penicillin and cephalosporin group of antibiotics. The parental β–lactamase gene appears to have originated from environmental microbes. However, bacteria acquired the resistance to newer β–lactam antibiotics by a series of point mutations within the lactamase gene. Such mutations are common in members of Enterobacteriaceae. Penicillin binding protein (PBP) is responsible for binding with β−lactam antibiotics and inhibition of cell wall synthesis. A mutational change in the mecA gene brings an alternative PBP (PBP2a) which in turn leads for methicillin resistance in S. aureus.
Horizontal Gene Transfer
Genetic elements like plasmid, transposon and integrons carry the antibiotic resistance genes (Boucher et al., 2007). These elements act as vectors and transfer resistance genes to other bacteria belonging to the members of the same species, or to another species or even a different genus (Dzidic and Bedekovic, 2003). Transformation, conjugation and transduction are the three different mechanisms in bacteria for horizontal gene transfer (HGT). The mechanism of HGT transfer in various antibiotics group has been summarized in Table 5.
Transformation involves the uptake of short naked DNA fragments and their homologous recombination in naturally competent bacteria. This is commonly observed in the species of streptococci, meningococci, Acinetobacter etc. In the recent years, Acinetobacter and other environmental genera have been associated to transmit the resistance genes from environmental microbes to clinical pathogens (Riesenfeld et al., 2004).
Conjugation involves the cell to cell contact via sexual pilli to transfer the piece of DNA. Sex pilli is formed by the responsible genes which is present only in the donor bacteria. Ultimately, the piece of DNA fragments having the resistance genes is transferred from resistant donors to previously susceptible bacteria. Recent studies have demonstrated that gut microbiota of human and animals frequently transmit the diverse ranges of antimicrobial resistance genes through conjugation.
Transduction involves the transfer of DNA from one bacterium into another via bacterial viruses called bacteriophage. It is less commonly linked with the transfer of antibiotic resistance genes compared to transformation and conjugation. However, phages are frequently associated with the formation of mobile genetic elements encoding the resistance and virulence genes.
Comparative analysis of plasmid from pre–antibiotic to the post–antibiotic era suggests that antibiotics resistance genes were present even in the pre–antibiotic era (Hughes and Datta, 1983). But the emergence of multi–resistance plasmids among the pathogens probably happened in the past five decades as a pressure of antibiotics use. The integron, a mobile DNA element, has two conserved segments flanking a central region carrying the antibiotic resistance gene. The integrons and their site–specific recombinase (integrase) enzyme are associated with the formation of multiple resistance plasmid and transposon.
Source of Antibiotics Resistance Genes
Evolution of antibiotics resistance genes in bacterial pathogens is a complex process. Growing number of evidence suggests that environment has reserve pool of antibiotic resistance genes (Martínez, 2008). Most of the antibiotic resistance genes acquired through horizontal gene transfer are expected to have originated from environmental microbes. Along with this various anthropogenic activities and evolution of existing bacterial enzymes are considered as source of resistance genes.
Environmental Microbes
Role of environmental microbes in spreading antibiotics resistance has been extensively explored (Riesenfeld et al., 2004; Allen et al., 2010). Most antibiotics used for treating infection are produced by environmental microbes. Antibiotic–producing bacteria protect themselves from the lethal effect of antibiotics through modification of targets and pumping it out through efflux pump as soon as it is formed. Discovery of these pathways in Streptomycetes led to the assumption that antibiotic resistance genes present in pathogenic bacteria have origin from antibiotic producing soil microbes. The quantification of antimicrobial resistance genes in antibiotic–producing bacterial strains reflects that a significant number of these strains are naturally multidrug resistant (D’Costa et al., 2006). Further, environmental microbes have been evaluated for their capacity to use antibiotics as a sole carbon and nitrogen source. Many of the bacterial strains belonging to proteobacteria, Burkholderia spp. and pseudomonads grow well on aminoglycosides, fluoroquinolones, and other classes of antimicrobials (Dantas et al., 2008). Streptomycetes produce a variety of β–lactamases. This is supposed to be the source of β–lactam resistance in clinical isolates (Kumarasamy et al., 2010). The environmental Kluyvera species have been found to be the origins of the CTX–M genes (Cantón and Coque, 2006). The quantum of data is sufficient to assume that the most potential pool of antibiotics resistance genes lies among the environmental microbes (Walsh, 2013).
Antibiotics Application in Agriculture
Apart from its therapeutic application in human and animals, antibiotics are widely used in agriculture, aquaculture, poultry industry and farm animals (Alderman and Hastings, 1998; Teuber, 2001; Wegener, 2003; Dibner and Richards, 2005; Cabello, 2006; Kemper, 2008; Li et al., 2013b). All these facilitate a vast reserve for antibiotic resistance genes in the natural habitat. The most commonly used antibiotic in agriculture is streptomycin and oxytetracycline (McManus et al., 2002). The excessive application of streptomycin resulted in development of resistant strains of Erwinia amylovora, Pseudomonas spp. and Xanthomonas campestris. Avoparcin is a glycopeptide antibiotic. It has been extensively used as a growth promoter in poultry and pig farms. This has resulted to development of resistance against vancomycin, one of the drugs of last resort to treat human infections (Bager et al., 1997). Therefore, it is necessary to curb the excessive use of antibiotics to prevent the spread of antibiotic resistance.
Anthropogenic Activity
Waste disposal and wastewater treatment plants are rich reservoirs for antimicrobial resistance genes and resistant microbes (Fick et al., 2009). This facilitates the horizontal gene transfer from these sources to human pathogens. This demands a better control of antibiotic release and environmental disposal. Recent work suggests that river water contaminated with quinolones enriches the qnr genes in water–borne bacteria and might be responsible for the transfer of qnr genes to pathogenic bacteria (Hirsch et al., 1999).
Bacterial Housekeeping Genes
Housing keeping genes having role in protection from antimicrobials including antibiotics in environmental bacteria might be the need to survive because soil, water and terrestrial lives produces variety of challenges. From there, the pathogenic and commensal microbes may also acquire genes of their need. In recent past several studies on herbal–drug resistance in pathogenic bacteria have revealed that it is not only use of antimicrobials which might be associated with existence/ emergence of drug resistance, microbes may have resistance to several types of herbal drugs which might have never been used in therapeutics (Singh et al., 2013). This phenomenon of resistance to non–use antimicrobials can easily be explained on the basis of presence of one or other house–keeping gene. Some of the bacterial housekeeping genes such as the sugar kinases and acyltransferases might have evolved to modify aminoglycoside antibiotics with the use of antimicrobials too (Davies and Davies, 2010).
In conclusion, it can be said that multiplicity of antimicrobials is not more than the ways to microbes know and have ability to evolve to counter those and emergence of the multiple drug resistance is the percussion.
REFERENCES
Abigail HD, April KR, and Clifford CS (2011). Super toxins from a super bug: structure and function of Clostridium difficile toxins. Biochem. J. 436: 517–526.
http://dx.doi.org/10.1042/BJ20110106
PMid:21615333
Abraham EP and Chain E (1940). An enzyme from bacteria able to destroy penicillin. Rev. Infect. Dis. 10: 677–678.
Alanis AJ (2005). Resistance to antibiotics: are we in the post–antibiotic era? Arch. Med. Res. 36: 697–705.
http://dx.doi.org/10.1016/j.arcmed.2005.06.009
PMid:16216651
Alderman DJ and Hastings TS (1998). Antibiotic use in aquaculture: development of antibiotic resistance–potential for consumer health risks. Int. J. Food Sci. Technol. 33: 139–155.
http://dx.doi.org/10.1046/j.1365-2621.1998.3320139.x
Alekshun MN and Levy SB (2007). Molecular mechanisms of antibacterial multidrug resistance. Cell. 128: 1037–1050.
http://dx.doi.org/10.1016/j.cell.2007.03.004
PMid:17382878
Allen HK, Donato J, Wang HH, Cloud–Hansen KA, Davies J Handelsman J (2010). Call of the wild: antibiotic resistance genes in natural environments. Nature Rev. Microbiol. 8: 251–259.
http://dx.doi.org/10.1038/nrmicro2312
PMid:20190823
Andre and Bryskier MD (2005). Antimicrobial agents: antibacterials and antifungals. ASM Press. SBN–10: 1555812376
Arias CA and Murray BE (2009). Antibiotic–resistant bugs in the 21st century—a clinical super–challenge. New Eng. J. Med. 360: 439–443.
http://dx.doi.org/10.1056/NEJMp0804651
PMid:19179312
Bager F, Madsen M, Christensen J and Aarestrup FM (1997). Avoparcin used as a growth promoter is associated with the occurrence of vancomycin–resistant Enterococcus faecium on Danish poultry and pig farms. Prev. Vet. Med. 31: 95–112.
http://dx.doi.org/10.1016/S0167-5877(96)01119-1
Blanchard JS (1996). Molecular mechanisms of drug resistance in Mycobacterium tuberculosis. Annual Rev. biochem. 65: 215–239.
http://dx.doi.org/10.1146/annurev.bi.65.070196.001243
PMid:8811179
Boucher Y, Labbate M, Koenig JE and Stokes HW (2007). Integrons: mobilizable platforms that promote genetic diversity in bacteria. Trends Microbiol. 1: 301–309.
http://dx.doi.org/10.1016/j.tim.2007.05.004
PMid:17566739
Breidenstein E, de la Fuente–Nú-ez C and Hancock RE (2011). Pseudomonas aeruginosa: all roads lead to resistance. Trends Microbiol. 19: 419–426.
http://dx.doi.org/10.1016/j.tim.2011.04.005
PMid:21664819
Brötz–Oesterhelt H and Brunner NA (2008). How many modes of action should an antibiotic have? Current Opinion Pharmacol. 8: 564–573.
http://dx.doi.org/10.1016/j.coph.2008.06.008
PMid:18621146
Cabello FC (2006). Heavy use of prophylactic antibiotics in aquaculture: a growing problem for human and animal health and for the environment. Environ. Microbiol. 8: 1137–1144.
http://dx.doi.org/10.1111/j.1462-2920.2006.01054.x
PMid:16817922
Cantón R and Coque TM (2006). The CTX–M β–lactamase pandemic. Current Opinion Microbiol. 9: 466–475.
http://dx.doi.org/10.1016/j.mib.2006.08.011
PMid:16942899
Dantas G, Sommer MO, Oluwasegun RD and Church GM (2008). Bacteria subsisting on antibiotics. Sci. 320: 100–103.
http://dx.doi.org/10.1126/science.1155157
PMid:18388292
Davies J (1994). Inactivation of antibiotics and the dissemination of resistance genes. Sci. 264: 375–382.
http://dx.doi.org/10.1126/science.8153624
Davies J and Davies D (2010). Origins and evolution of antibiotic resistance. Microbiology and Mol. Biol. Rev. 74: 417–433.
http://dx.doi.org/10.1128/MMBR.00016-10
PMid:20805405 PMCid:PMC2937522
D'Costa VM, McGrann KM, Hughes DW and Wright GD (2006). Sampling the antibiotic resistome. Sci. 311: 374–377.
http://dx.doi.org/10.1126/science.1120800
PMid:16424339
Dhama K, Chakraborty S, Mahima, Wani MY, Verma AK, Deb R, Tiwari R and Kapoor S (2013a). Novel and emerging therapies safeguarding health of humans and their companion animals: A review. Pak. J. Bio. Sci. 16(3): 101–111. doi: 10.3923/pjbs.2013.101.111
http://dx.doi.org/10.3923/pjbs.2013.101.111
Dhama K, Chakraborty S, Wani MY, Tiwari R and Barathidasan R (2013b). Cytokine therapy for combating animal and human diseases – A review. Res. Opin. Anim. Vet. Sci. 3(7): 195–208.
Dibner JJ and Richards JD (2005). Antibiotic growth promoters in agriculture: history and mode of action. Poul. Sci. 84: 634–643.
http://dx.doi.org/10.1093/ps/84.4.634
PMid:15844822
Dzidic S and Bedekovic V (2003). Horizontal gene transfer–emerging multidrug resistance in hospital bacteria. Acta Pharmacol. Sin. 24: 519–526.
PMid:12791177
Fajardo A, Martinez–Martin N, Mercadillo M, Galan JC, Ghysels B, Matthijs S and Martinez JL (2008). The neglected intrinsic resistome of bacterial pathogens. PloS One. 3: e1619.
Fick J, Söderstrom H, Lindberg RH, Phan C, Tysklind M and Larsson DG (2009). Contamination of surface, ground, and drinking water from pharmaceutical production. Environ. Toxicol. Chem. 28: 2522–2527.
http://dx.doi.org/10.1897/09-073.1
PMid:19449981
Foster TJ (2004). The Staphylococcus aureus "superbug". J. Clin. Invest. 114: 1693–1696.
http://dx.doi.org/10.1172/JCI23825
http://dx.doi.org/10.1172/JCI200423825
PMid:15599392 PMCid:PMC535074
Hancock RE (1998). Resistance mechanisms in Pseudomonas aeruginosa and other nonfermentative gram–negative bacteria. Clin. Infect. Dis. 27:(Supplement 1) S93–S99.
http://dx.doi.org/10.1086/514909
PMid:9710677
Hawkey PM (1998). The origins and molecular basis of antibiotic resistance. B.M.J. 317: 657–660.
http://dx.doi.org/10.1136/bmj.317.7159.657
PMid:9727999 PMCid:PMC1113838
Hazbon MH, Brimacombe M, del Valle MB, Cavatore M, Guerrero MI, Varma–Basil M and Alland D (2006). Population genetics study of isoniazid resistance mutations and evolution of multidrug–resistant Mycobacterium tuberculosis. Antimicro. Agents Chemother. 50: 2640–2649.
http://dx.doi.org/10.1128/AAC.00112-06
PMid:16870753 PMCid:PMC1538650
Higgins CF (2007). Multiple molecular mechanisms for multidrug resistance transporters. Nature. 446: 749–757.
http://dx.doi.org/10.1038/nature05630
PMid:17429392
Hirsch R, Ternes T, Haberer K and Kratz KL (1999). Occurrence of antibiotics in the aquatic environment. Sci. Total Environ. 225: 109–118.
http://dx.doi.org/10.1016/S0048-9697(98)00337-4
Hooper DC (2001). Emerging mechanisms of fluoroquinolone resistance. Emerg. Infect. Dis. 7: 337–341.
http://dx.doi.org/10.3201/eid0702.010239
PMid:11294736 PMCid:PMC2631735
Howden BP, Davies JK, Johnson PD, Stinear TP and Grayson ML (2010). Reduced vancomycin susceptibility in Staphylococcus aureus, including vancomycin–intermediate and heterogeneous vancomycin–intermediate strains: resistance mechanisms, laboratory detection, and clinical implications. Clin. Microbiol. Rev. 23: 99–139.
http://dx.doi.org/10.1128/CMR.00042-09
PMid:20065327 PMCid:PMC2806658
Hughes VM, and Datta N (1983). Conjugative plasmids in bacteria of the 'pre–antibiotic'era. Nature 302: 725–726.
http://dx.doi.org/10.1038/302725a0
PMid:6835408
Jana S and Deb JK (2006). Molecular understanding of aminoglycoside action and resistance. Appl. Microbiol. Biotechnol. 70: 140–150.
http://dx.doi.org/10.1007/s00253-005-0279-0
PMid:16391922
Kemper N (2008). Veterinary antibiotics in the aquatic and terrestrial environment. Ecol. Indicat. 8: 1–13.
http://dx.doi.org/10.1016/j.ecolind.2007.06.002
Kumarasamy KK, Toleman MA, Walsh TR, Bagaria J, Butt F, Balakrishnan R and Woodford N (2010). Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and epidemiological study. The Lancet Infect. Dis. 10: 597–602.
http://dx.doi.org/10.1016/S1473-3099(10)70143-2
Laurenzo D and Mousa SA (2011). Mechanisms of drug resistance in Mycobacterium tuberculosis and current status of rapid molecular diagnostic testing. Acta. Tropica. 119: 5–10.
http://dx.doi.org/10.1016/j.actatropica.2011.04.008
PMid:21515239
Levy SB and Marshall B (2004). Antibacterial resistance worldwide: causes, challenges and responses. Nat. Med. 10: S122–S129.
http://dx.doi.org/10.1038/nm1145
PMid:15577930
Li W, Atkinson GC, Thakor NS, Allas U, Lu CC, Chan KY, Tenson T, Schulten K, Wilson KS, Hauryliuk V and Frank J (2013a). Mechanism of tetracycline resistance by ribosomal protection protein Tet (O). Nature Commun. 4: 1477. doi: 10.1038/ncomms2470.
http://dx.doi.org/10.1038/ncomms2470
Li YX, Zhang XL, Li W, Lu XF, Liu B and Wang J (2013b). The residues and environmental risks of multiple veterinary antibiotics in animal faeces. Environ. Monitor. Assess. 185: 2211–2220.
http://dx.doi.org/10.1007/s10661-012-2702-1
PMid:22692716
Lindsay JA and Holden MT (2004). Staphylococcus aureus: superbug, super genome? Trend. Microbiol. 12: 378–385.
http://dx.doi.org/10.1016/j.tim.2004.06.004
PMid:15276614
Livermore DM (2002). Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: our worst nightmare? Clin. Infecti. Dis. 34: 634–640.
http://dx.doi.org/10.1086/338782
PMid:11823954
Mah TFC and O'Toole GA (2001). Mechanisms of biofilm resistance to antimicrobial agents. Trend. Microbiol. 9: 34–39.
http://dx.doi.org/10.1016/S0966-842X(00)01913-2
Marshall BM and Levy SB (2011). Food animals and antimicrobials: impacts on human health. Clinicl. Microbiol. Rev. 24: 718–733.
http://dx.doi.org/10.1128/CMR.00002-11
PMid:21976606 PMCid:PMC3194830
Martinez JL (2008). Antibiotics and antibiotic resistance genes in natural environments. Sci. 321:, 365–367.
http://dx.doi.org/10.1126/science.1159483
PMid:18635792
McManus PS, Stockwell VO, Sundin GW and Jones AL (2002). Antibiotic use in plant agriculture. Ann. Rev. Phytopathol. 40: 443–465.
http://dx.doi.org/10.1146/annurev.phyto.40.120301.093927
PMid:12147767
Piddock LJ (2006). Multidrug–resistance efflux pumps? Not just for resistance. Nat. Rev. Microbiol. 4: 629–636.
http://dx.doi.org/10.1038/nrmicro1464
PMid:16845433
Poirel L and Nordmann P (2006). Carbapenem resistance in Acinetobacter baumannii: mechanisms and epidemiology. Clin. Microbiol. Infect. 12: 826–836.
http://dx.doi.org/10.1111/j.1469-0691.2006.01456.x
PMid:16882287
Poole K (2005). Efflux–mediated antimicrobial resistance. J. Antimicrob. Chemother. 56: 20–51.
http://dx.doi.org/10.1093/jac/dki171
PMid:15914491
Rattan A, Kalia A and Ahmad N (1998). Multidrug–resistant Mycobacterium tuberculosis: molecular perspectives. Emerg. Infect. Dis. 4: 195–209.
http://dx.doi.org/10.3201/eid0402.980207
PMid:9621190 PMCid:PMC2640153
Riesenfeld CS, Goodman RM and Handelsman J (2004). Uncultured soil bacteria are a reservoir of new antibiotic resistance genes. Environ. Microbiol. 6: 981–989.
http://dx.doi.org/10.1111/j.1462-2920.2004.00664.x
PMid:15305923
Rodríguez–Verdugo A, Gaut BS and Tenaillon O (2013). Evolution of Escherichia coli rifampicin resistance in an antibiotic–free environment during thermal stress. BMC Evol. Biol., 13: 50
http://dx.doi.org/10.1186/1471-2148-13-50
PMid:23433244 PMCid:PMC3598500
Samanidou VF and Evaggelopoulou EN (2008). Chromatographic analysis of banned antibacterial growth promoters in animal feed. J. Sep. Sci. 31: 2091–2112.
http://dx.doi.org/10.1002/jssc.200800075
PMid:18491356
Schwarz S and Chaslus–Dancla E (2001). Use of antimicrobials in veterinary medicine and mechanisms of resistance. Vet. Res. 32: 201–225.
http://dx.doi.org/10.1051/vetres:2001120
PMid:11432414
Schwarz S, Kehrenberg C and Walsh TR (2001). Use of antimicrobial agents in veterinary medicine and food animal production. Int. J. Antimicrol. Agent. 17: 431–437.
http://dx.doi.org/10.1016/S0924-8579(01)00297-7
Singh BR (2011). Drug resistant bacteria in the environment: In: Environmental Security, SR Garg (ed), IBDC Publishers, Lucknow, India, pp.313–326.
Singh BR (2013). Antimicrobial sensitivity assay and antimicrobial chemotherapy in Animals: A Practical approach. In: Diseases of Animals: Diagnosis and Management, BR Singh and R Somvanshi (eds.), Indian Veterinary Research Institute, Izatnagar, bareilly, pp.7–31.
Singh BR, Singh V, Ebibeni N and Singh RK (2013). Antimicrobial and herbal drug resistance in enteric bacteria isolated from faecal droppings of common house lizard/gecko (Hemidactylus frenatus). Int. J. Microbiol. 2013: 8 pages, doi:10.1155/2013/340848.
http://dx.doi.org/10.1155/2013/340848
Spratt BG (1994). Resistance to antibiotics mediated by target alterations. Sci. 264: 388–393.
http://dx.doi.org/10.1126/science.8153626
Teuber M (2001). Veterinary use and antibiotic resistance. Current Opinion Microbiol. 4: 493–499.
http://dx.doi.org/10.1016/S1369-5274(00)00241-1
Tiwari R, Chakraborty S, Dhama K, Rajagunalan S, Singh SV (2013a). Antibiotic resistance – an emerging health problem: causes, worries, challenges and solutions – a review. Int. J. Curr. Res. 5(07): 1880–1892.
Tiwari R, Dhama K, Chakraborty S, Kumar A, Rahal A and Kapoor S (2013b). Bacteriophage therapy for safeguarding animal and human health: A review. Pak. J. Biol. Sci. doi: 10.3923/pjbs.2013.
Walsh C (2000). Molecular mechanisms that confer antibacterial drug resistance. Nature. 406: 775–781.
http://dx.doi.org/10.1038/35021219
PMid:10963607
Walsh TR and Howe RA (2002). The prevalence and mechanisms of vancomycin resistance in Staphylococcus aureus. Ann. Rev. Microbiol. 56: 657–675.
http://dx.doi.org/10.1146/annurev.micro.56.012302.160806
PMid:12142482
Walsh F (2013). Investigating antibiotic resistance in non–clinical environments. Front. Microbiol. 4: 19.
http://dx.doi.org/10.3389/fmicb.2013.00019
PMid:23423602 PMCid:PMC3573686
Wegener HC (2003). Antibiotics in animal feed and their role in resistance development. Current Opinion Microbiol. 6: 439–445.
http://dx.doi.org/10.1016/j.mib.2003.09.009
PMid:14572534
WHO (2013). Antimicrobial resistance. www.who.int.
Wright GD (2005). Bacterial resistance to antibiotics: enzymatic degradation and modification. Adv. Drug Del. Rev. 57: 1451–1470
http://dx.doi.org/10.1016/j.addr.2005.04.002
PMid:15950313

