
Heritability Estimates and Correlation Analysis for Production Traits in Fcv Tobacco




Research Article
Heritability Estimates and Correlation Analysis for Production Traits in Fcv Tobacco
Sheraz Ahmed* and Fida Mohammad
Department of Plant Breeding and Genetics, The University of Agriculture, Peshawar, 25130, Khyber Pakhtunkhwa, Pakistan.
Abstract | Knowledge of heritability is an essential prerequisite for any crop improvement program. One hundred recombinant inbred lines (RILs) derived from four F4 populations of Flue Cured Virginia (FCV) tobacco were tested to identify effective selection indices. Experimental material was planted at two locations i.e. Mardan (E-1, E-3 and E-5) and Mansehra (E-2, E-4 and E-6) using alpha lattice design with three replicates during 2012/13, 2013/14 and 2014/15. Heritability in broad sense was generally low for all traits except nicotine and reducing sugar. Days to flowering was the most environment responsive trait and its heritability fluctuated between 0.22 and 0.91. Plant height was significantly associated with yield at phenotypic level only. Yield exhibited significantly negative phenotypic correlation with days to flowering. Similarly, yield was positively correlated with leaves per plant, green leaves weight per plot, cured leaves weight per plot and grade index at both phenotypic and genotypic level. Based on findings of the present study, selection among RILs would be more effective on the basis of nicotine, reducing sugar, leaves per plant, green leaves weight, cured leaves weight and grade index.
Received | March 01, 2016; Accepted | March 25, 2016; Published | April 15, 2017
*Correspondence | Sheraz Ahmed, Department of Plant Breeding and Genetics, The University of Agriculture, Peshawar, 25130, Khyber Pakhtunkhwa, Pakistan; Email: sheraazahmed@yahoo.com
Citation | Ahmed, S. and F. Mohammad. 2017. Heritability estimates and correlation analysis for production traits in fcv tobacco. Sarhad Journal of Agriculture, 33(2): 212-219.
DOI | http://dx.doi.org/10.17582/journal.sja/2017/33.2.212.219
Keywords | FCV tobacco, Nicotiana tabacum, Heritability, Genetic correlation, Crop improvement
Introduction
Tobacco belongs to the genus Nicotiana of the nightshade family, Solanaceae. Other members of this family are potatoes (Solanum tuberosum L.) and tomatoes (Solanum lycopersicum L.). This family includes plants from herbs to trees as well as a number of important agricultural crops, medicinal plants, spices and weeds, of which a considerable number is of major economic importance as field crops, vegetables or ornamental species throughout the world (Darvishzadeha and Maleki, 2012). Nicotiana genus also called as a genus of herbaceous plants and shrubs has more than 70 different species indigenous to the Americas, Australia, South West Africa and the South Pacific. But only two species of Nicotiana (N. tabacum L. and N. rustica L.) are grown worldwide for production of tobacco leaf for cigarettes and other tobacco products manufactured from dried tobacco leaves, including cigars, snuff, pipe tobacco, chewing tobacco and flavoured shisha (Taj, 1994). The flue-cured tobacco (Nicotiana tabacum L.) in particular is very important for tobacco industry in the world (Zhang et al., 2008).
Tobacco is an important cash crop in numerous countries. Likewise in Pakistan, despite cultivation on limited irrigated area (0.27%), tobacco is a great source of revenue, foreign exchange and employment. The contribution of tobacco to the federal exchequer is over Rs. 110 billion which is more than any other crop. In addition, export of raw tobacco and its products is earning handsome amount of 24 million US$ (PTB, 2016). Tobacco industry is
Table 1: List of parent cultivars (checks) and RILs with parentage.
| S/n | Code | Variety/line | Parentage | Main features |
| 1 | Chk1 | Speight G-28 | (Coker-139 × Oxford 1-181) and NC-95 | Recommended variety for several decades in Pakistan, having modest yield and quality. Plants are shorter than many varieties which bear 25 leaves per plant. Medium to late maturity. |
| 2 | Chk2 | Speight G-126 | K-326 × Speight G-96 | Moderate yield with inferior cured leaf quality. Late maturing than most of the varieties Good holding ability. |
| 3 | Chk3 | NC-606 | NC-729 × NC-82 | Produces 30 good quality leaves per plant. Taller plants with longer internodal length. |
| G1 – G100 | G1 – G25 | Spt G-28 × Spt G-126 | Segregating populations were advanced in bulk till F4 generation. Single plant selection in F4 generation was made under rainfed condition. | |
| G26 – G50 | Spt G-126 × Spt G-28 | |||
| G51 – G75 | Spt G-28 × NC-606 | |||
| G76 – G100 |
Spt G-126 × NC-606 |
tbasically capital inensive and only tobacco processing and manufacturing units require human resource. However, tobacco farming is still very labour intensive. Currently, about 80 thousand people are involved in its cultivation, 50 thousand are engaged in 21 cigarettes manufacturing companies while another one million find indirect employment (PTB, 2016). However, due to lack of considerable efforts in tobacco breeding, no indigenous tobacco cultivar is in commercial cultivation. As a result, cultivation of susceptible tobacco varieties with modest yield is being in practice. Presently, the country is relying on introduced genetic material for commercial cultivation of tobacco. This necessitated the need of native tobacco breeding program to alleviate the problems confronting tobacco industry in Pakistan.
High productivity is the ultimate goal of every breeding program. The knowledge regarding the nature of association among various plant characters is crucial for yield improvement. Most of the plant traits are quantitative in nature. Likewise, yield is one of them which is the result of many associations and interactions of component characters. It is widely accepted among researchers that the structure and nature of yield could better be understood by examining its components traits. This helps plant breeders to introgress desirable alleles in breeding populations to realize the goal of high productivity. Similarly, effective selection is the backbone of any breeding plan. Heritability predicts the behaviour and interaction of genes in succeeding generations thereby, providing appropriate selection criteria. High heritability indicates more improvement in lesser time period. Many researchers have advocated the use of genetic advance with heritability as heritability alone cannot provide enough information regarding the pattern of gene actions. High magnitude of heritability coupled with genetic advance provides clue about the presence of fixable (additive) genes which ease and increase the efficacy of selection in cultivar development (Shukla et al., 2004). Contrarily, high heritability with low genetic advance suggests the presence of non-fixable (dominant/epistatic) genes which could be exploited through heterosis breeding (Shoukat et al., 2007). Nevertheless, insufficient information in FCV tobacco is currently available in this regard. Therefore, this experiment was carried out to; i) establish association among yield related traits in FCV tobacco for yield improvement and ii) estimate heritability in broad sense and genetic advance over environments.
Materials and Methods
Plant material and experimental sites
This study was conducted in the two experimental stations of Pakistan Tobacco Board. Plant material comprising 100 RILs of FCV tobacco was planted using alpha lattice design in three replicates at the Tobacco Research Station, Mardan (Plain) and Tobacco Research Station, Mansehra (Hilly) during 2012/13, 2013/14 and 2014/15 (3 years). These RILs were derived from four F4 populations of Speight G-28/Speight G-126, Speight G-126/Speight G-28, Speight G-28/NC-606 and Speight G-126/ NC-606 (Table 1). Locations in each year i.e. Mardan 2012/13, Mansehra 2012/13, Mardan 2013/14, Mansehra 2013/14, Mardan 2014/15 and Mansehra 2014/15 were referred to as six environments E-1, E-2, E-3, E-4, E-5 and E-6, respectively.
DTF: days to flowering; PH: plant height; LP: leaves plant-1; IL: internodal length; LA: leaf area; GW: green leaves weight plot-1; GL: number of green leaves kg-1; CW: cured leaves weight plot-1; CL: number of cured leaves kg-1; GI: grade index, Nic: nicotine; RS: reducing sugars
Seedbed preparation and nursery raising
Tobacco seeds are very small in size and hence are prone to injuries in fields. Therefore, seedlings are usually raised on nursery beds in mid-December. Seedbeds were raised 15 cm above the ground level measuring 10 m2 (10m × 1 m). Soil in seedbeds was cleaned from all kind of weeds and other plant residues. Farm yard manure was applied on the surface of seedbeds to improve water holding capacity. About 1 g of seed was mixed with dry fine sand and then broadcasted evenly over the seedbed and immediately watered. Seedbeds were watered and weeded on regular basis. Seedlings having 4 to 5 leaves were then transplanted in mid-March at Mardan while about two weeks earlier at Mansehra.
Statistical analysis
Field data were subjected to analysis of variance using appropriate model for alpha lattice design following Steel et al. (1996). Upon significant genotype by environment (GE) interaction, analysis of variance at each environment was also carried out. Phenotypic and genotypic correlation coefficients were calculated following procedures of Singh and Chaudhary (1985). Data for each environment were statistically analysed using appropriate model for alpha lattice design. Upon significant variation among genotypes, broad sense heritability was calculated from ANOVA for each trait by calculating variance components according to Marwede et al. (2004) as under;
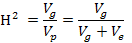
Where:
Vg: genetic variance; Vp: phenotypic variance; Ve: error variance.
Genetic advance was calculated following Allard’s (1964) formula;
Where:
i: 1.76 for 10% selection intensity; H2: broad sense heritability of a trait x; Vp: phenotypic variance of a trait x.
Similarly, genetic advance as percentage mean was calculated as;
Where:
X: grand mean for a trait x; GA: genetic advance.Results and Discussion
Heritability and genetic advance
Field data for various traits recorded in each environment were subjected to analysis of variance. Analysis revealed significant differences among RILs for some traits while non-significant for others (Table 2). Heritability and genetic advance were estimated only for traits where RILs exhibited genetic differences.
Table 3: Heritability estimates* and genetic advance for various traits of 100 RILs of FCV tobacco evaluated across six environments.
| Traits | E1 | E2 | E3 | E4 | E5 | E6 | |
| DTF |
H2 |
0.90 | - | 0.92 | 0.22 | 0.61 |
0.25 |
| GA | 6.91 | - | 7.24 | 1.10 | 4.36 | 0.97 | |
| GA% | 11.2 | - | 11.7 | 1.5 | 6.9 | 1.4 | |
| PH |
H2 |
0.25 | 0.10 | 0.24 | - | - | 0.14 |
| GA | 4.92 | 1.11 | 3.76 | - | - | 1.11 | |
| GA% | 4.7 | 1.4 | 3.2 | - | - | 1.2 | |
| IL |
H2 |
0.41 | - | - | - | 0.12 | - |
| GA | 0.43 | - | - | - | 0.26 |
- |
|
| GA% | 8.3 | - | - | - | 4.4 | - | |
| LA |
H2 |
0.29 | 0.10 | 0.45 | - | - | - |
| GA | 62.78 | 13.85 | 102.95 | - | - | - | |
| GA% | 8.0 | 2.1 | 11.8 | - | - | - | |
| LP |
H2 |
0.51 | - | 0.10 | - | 0.13 | - |
| GA | 2.81 | - | 0.30 | - | 0.61 | - | |
| GA% | 13.9 | - | 1.5 | - | 3.6 |
- |
|
| GP |
H2 |
0.35 | 0.31 | 0.22 | 0.12 | - | 0.14 |
| GA | 0.82 | 0.92 | 0.42 | 0.25 | - | 0.44 | |
| GA% | 9.3 | 13.8 | 7.4 | 3.8 | - | 6.9 | |
| GL |
H2 |
0.35 | 0.37 | 0.17 | 0.14 | - | 0.14 |
| GA | 2.04 | 3.55 | 1.49 | 0.92 | - | 1.18 | |
| GA% | 9.3 | 12.6 | 6.1 | 3.5 | - | 6.2 | |
| CP |
H2 |
0.25 | 0.13 | 0.21 | - | - | - |
| GA | 0.1 | 0.06 | 0.08 | - | - | - | |
| GA% | 7.3 | 4.8 | 7.2 | - | - | - | |
| Yield |
H2 |
0.25 | 0.13 | 0.21 | - | - | - |
| GA | 175.8 | 115.1 | 142.1 | - | - | - | |
| GA% | 7.3 | 4.8 | 7.2 | - | - | - | |
| Nic |
H2 |
0.88 | 0.92 | 0.98 | 0.97 | 0.95 | 0.78 |
| GA | 0.41 | 0.18 | 0.75 | 0.67 | 0.61 | 0.39 | |
| GA% | 22.0 | 11.3 | 31.2 | 28.4 | 23.0 | 17.8 | |
| RS |
H2 |
0.98 | 0.94 | 0.67 | 0.98 | 0.95 |
0.91 |
| GA | 3.31 | 6.08 | 4.5 | 3.84 | 3.33 | 1.98 | |
| GA% | 26.7 | 40.4 | 25.4 | 24.7 | 23.9 |
15.0 |
DTF: days to flowering; PH: plant height; LP: leaves plant-1; IL: internodal length; LA: leaf area; GW: green leaves weight plot-1; GL: number of green leaves kg-1; CW: cured leaves weight plot-1; CL: number of cured leaves kg-1; GI: grade index, Nic: nicotine; RS: reducing sugars
* Heritability in broad sense was estimated for traits where differences were significant among genotypes.
High heritability with moderate to high genetic advance was estimated for days to flowering at E-1 (0.90) and E-3 (0.92) (Mardan) whereas, at E-4 and E-6 (Mansehra) heritability dropped to 0.22 and 0.25, respectively (Table 3). Low heritability estimates are indication of unfavourable environmental conditions for days to flowering at Mansehra. This signalled slow progress through selection for days to flowering at Mansehra. Conversely, higher estimates of heritability accompanied by high genetic advance at Mardan indicated that the inheritance of days to flowering is relatively simple and governed by the additive type of gene action. Thus, selection for days to flowering in FCV tobacco would be more suitable at Mardan and significant achievement is expected through early generation cycles. Similarly, heritability estimates for plant height, leaf area and leaves plant-1 were lower at Mansehra than Mardan (Table 3). This suggested that these traits were environment-specific and thus gain from selection would be more at later generations. The major role of environmental effects on the phenotypic expression of number of leaves plant-1 and leaf area have been previously reported by Sadeghi et al. (2009). Low to moderate heritability with modest genetic advance was estimated for green leaves weight plot-1, green leaves kg-1 and cured leaves weight plot-1 at all environments. Lower heritability estimate insinuated the sluggish progress of these traits over generations. Thus, selection on phenotypic performance is not applicable due to masking effect of environment on genotypic expression. Heritability estimated for yield fluctuated between 0.13 and 0.25 with decent genetic advance (Table 3). This suggested the presence of higher environmental noise which could be an indication towards careful selection of this trait at later generations. Current results confirmed the finding of Sadeghi et al. (2009) that the phenotypic expression of dry leaf yield was dominated by environmental effects. Contrary to other traits, high heritability accompanied by high genetic advance was estimated for nicotine and reducing sugar contents at all environments. This indicated that the genetic effects in the phenotypic expression of these traits were of additive in nature, therefore earlier selection would be effective. Similar results were also reported by Katba (2012) and Nama (2011). Results of this study were contradictory to the findings of Kara and Esendal (1995) who reported non-additive type of gene action for nicotine and reducing sugar contents and favored selection at later stage for these traits in FCV tobacco. Inconsistency in results could be attributed to differences in test environments or genotypes or may be both.
DTF: days to flowering; PH: plant height; LP: leaves plant-1; IL: internodal length; LA: leaf area; GW: green leaves weight plot-1; GL: number of green leaves kg-1; CW: cured leaves weight plot-1; CL: number of cured leaves kg-1; GI: grade index, Nic: nicotine; RS: reducing sugars. *, **: Significant at P < 0.05 and P < 0.01, respectively
Overall, high heritability coupled with high genetic advance for nicotine and reducing sugar was estimated at all environments suggesting the consistent involvement of fixable genes in the inheritance of these traits. This also suggested that these traits were least affected by environmental variances. Contrarily, heritability for days to flowering fluctuated between 0.22 and 0.92 across environments. This suggested that phenotypic expression of this trait was forced by environments in specific direction. Performance of genotypes for yield and yield components like green leaves weight plot-1 and cured leaves weight plot-1 were also dictated by fluctuating environments. Relative changes in heritability over environments may be attributed to the presence of allelic differences and perhaps different loci might have expressed under different environmental conditions (Eid, 2009). Generally, based on the estimates of heritability in the current study, traits other than nicotine and reducing sugar would require careful selection at later generations.
Phenotypic and genotypic correlation coefficients
Data for various morpho-agronomic traits of 100 RILs were recorded at Mardan and Mansehra across three years. Correlation coefficients calculated on phenotypic and genotypic data at both locations were referred to as phenotypic and genotypic correlations, respectively. Yield had significant associations with various traits over both locations. Details of results are given as under;
Correlation coefficients at Mardan (combined over three years): Magnitude of genetic correlations was higher for some traits than their corresponding phenotypic correlations suggesting the preponderance of genetic coherence in the expression of respective traits. On the other hand, higher magnitude of phenotypic correlations indicated that the association between two traits was environment-specific. Correlation analysis unveiled significantly positive associations of yield with plant height (rp= 0.078), however, non-significant association was detected at genotypic level (Table 4). This was contrary to the findings of Ahmed et al. (2014) who reported significantly positive phenotypic correlation of yield with plant height in FCV tobacco. Difference in findings may be mainly due to the environmental difference but to some extent it may be due to variation in germplasm. Significantly positive relationship of yield was also observed with leaves plant-1 (rp= 0.608, rg= 0.584), green leaves weight plot-1 (rp= 0.879, rg= 0.906) and grade index (rp= 0.790, rg= 0.807) at both phenotypic and genotypic levels (Table 4). This indicated higher contributing role of these traits towards realization of higher yield. Therefore, special attention may be given
DTF: days to flowering; PH: plant height; LP: leaves plant-1; IL: internodal length; LA: leaf area; GW: green leaves weight plot-1; GL: number of green leaves kg-1; CW: cured leaves weight plot-1; CL: number of cured leaves kg-1; GI: grade index, Nic: nicotine; RS: reducing sugars. *, **: Significant at P < 0.05 and P < 0.01, respectively
to leaves plant-1, green leaves weight plot-1 and grade index in selection program for higher productivity in FCV tobacco. Patel and Kinganokar (2005) and Katba (2012) also urged selection based on plant height and number of leaves plant-1 to isolate high yielding tobacco lines. Phenotypic and genotypic correlations of yield with cured leaves weight plot-1 were perfect (rg=rp= 1) which could be attributed to the fact that cured leaves weight plot-1 is an extrapolating factor for yield in FCV tobacco.
Significantly negative phenotypic correlation of yield was detected with days to flowering (rp= -0.095). This might be due to slow vegetative growth rate as a result of less favourable environmental conditions (Table 4). This indicated that it was not the length of biological life but the growing degree days responsible for realizing quantum of growth. Similarly, yield had significant negative correlations with internodal length (rp= -0.494, rg= -0.393), number of green leaves kg-1 (rp= -0.309, rg= -0.248) and number of cured leaves kg-1 (rp= -0.330, rg= -0.346) both at phenotypic and genotypic levels which showed that it was not the number of green leaves and cured leaves per kg but the size of leaves that realized higher yield. Higher number of leaves kg-1 would result in smaller leaves which happened to have significant reduction in tobacco yield. Datta (2002) also found significant contribution of leaves plant-1 and plant height towards higher yield in FCV tobacco.
Correlation coefficients at Mansehra (combined over three years): Higher values of genotypic correlation than their corresponding phenotypic correlations indicated the important role of genetics in the associations of various traits. Yield was significantly associated with days to flowering (rp= 0.114) at phenotypic level while significantly negative association was observed at genotypic level (rg= -0.265) (Table 5). Lower average temperature at Mansehra not only caused late flowering but also prolonged vegetative phase as compared to Mardan. The positive correlation of days to flowering with yield at Mansehra might be due to long vegetative phase thereby, offering more time for the plant to realize its maximum potential. That is why, the psitive association of days to flowering with yield was only reflected at phenotypic level. Hence, late flowering plants of FCV tobacco may be suitable at Mansehra only. Significantly positive correlations of yield were exhibited by plant height (rp= 0.220) and nicotine content (rp= 0.182) at phenotypic level only while no association was observed at genotypic level. This implies that the associations of these traits were environment-specific. Association between plant height and yield in tobacco has been reported by Dyulgerski and Radoukova (2014). Similarly, yield was also positively associated with leaves plant-1 (rp= 0.237, rg= 0.475), green leaves weight plot-1 (rp= 0.554, rg= 0.680), cured leaves weight plot-1 (rp= 1.000, rg= 1.000) and grade index (rp= 0.569, rg= 0.643) at both phenotypic and genotypic levels, confirming the true relationship of these traits with yield in FCV tobacco (Table 5). These traits could serve as desirable selection indices when making selection for higher yield in tobacco. The association of leaves plant-1 with yield has been extensively reported by earlier researchers such as Maleki et al. (2011), Bhumarannavar (2009), Patel and Makawana (2002), Lalithadevi et al. (2002) in FCV tobacco.
Yield was negatively associated with internodal length (rp= -0.068, rg= -0.334), green leaves kg-1 (rp= -0.515, rg= -0.575) and cured leaves kg-1 (rp= -0.624, rg= -0.620) at both phenotypic and genotypic levels. This suggests that these traits have negative impact on tobacco yield hence, smaller values of these traits are encouraged for higher yield in FCV tobacco.
Conclusions
Heritabilty in broad sense for various traits fluctuated across environments which indicated sporadic types of environmental changes. Generally, lower estimates of heritability for various traits signalled towards slow progress through selection. Phenotypic and genotypic correlation coefficients for yield with other traits revealed that leaves plant-1, green leaves weight plot-1, cured leaves weight plot-1 and grade index had major contributions towards higer yield in FCV tobacco. Therefore, these traits may be considered as selection criteria when making selection for higher yield in FCV tobacco.
Acknowledgment
Authors would like to thank Pakistan Tobacco Board for their logistic support during the course of this research.
Authors Contributions
This research article is an integral part of doctoral study of the first author Sheraz Ahmed (SA). SA and Fida Mohammad (FM) formulated the research. SA performed the experiments and wrote the first draft of the manuscript. FM made critical corrections in the first draft. All authors read and approved the final manuscript.
References
Allard, R. 1964. Principles of plant breeding. John Wiley and Sons. Inc. New York.
Bhumarannavar, K. 2009. Evaluation of different germplasm lines for alternative uses; disease and pest reaction in bidi tobacco. M. Sc. (Agri.) thesis submitted to University of Agricultural Sciences, Dharwad.
Datta, A.K. 2002. Study of morpho-physiological traits related to drought under irrigated and rainfed conditions in bidi tobacco (N. tabacum L.). M. Sc. (Agri.) thesis submitted to Gujarat Agricultural University, Sardarkrushinagar.
Darvishzadeh, R. and H.H. Maleki. 2012. Analysis of genetic variation for morphological and agronomic traits in Iranian oriental tobacco (Nicotiana tabacum L.) genotypes. Crop Breed. J. 2(1): 57-61.
Dyulgerski, Y.K. and T. Radoukova. 2014. Correlations between morphological and productive parameters in Burley tobacco. Agric. Sci. Technol. 6(2): 197-198
Eid, M.H. 2009. Estimation of heritability and genetic advance of yield traits in wheat (Triticum aestivum L.) under drought condition. Intl. J. Genet. Mol. Biol. 1(7): 115-120.
Kara, S.M. and E. Esendal. 1995. Heterosis and combining ability analysis of some quantitative characters in Turkish tobacco. Tob. Res. 21(1-2): 16-22.
Katba, P. 2012. Genetic variability, correlation and path analysis for yield in tobacco (Nicotiana rustica L.). M. Sc. (Agric.) thesis submitted to Anand Agricultural University, Anand.
Lalithadevi, D., R. Lakshminarayana and J.B. Atluri, 2002. Genetic variability and correlation studies on seed and other quantitative characters in tobacco (Nicotiana tabacum L.). Tob. Res. 28(2): 90-96.
Maleki, H.H., G. Karimzadeh, R. Darvishzadeh and A. Sarrafi. 2011. Correlation and sequential path analysis of some agronomic traits in tobacco (Nicotiana tabacum L.) to improve dry leaf yield. Aust. J. Crop Sci. 5(12): 1644-1648.
Marwede, V., A. Schierholt, C. Möllers and H.C. Becker. 2004. Genotype × environment interactions and heritability of tocopherol contents in canola. Crop Sci. 44: 728-731. https://doi.org/10.2135/cropsci2004.0728
Patel, A. D. and S.K. Kingaonkar. 2005. Genetic analysis in tobacco (Nicotiana tabacum L.). Tob. Res. 32(1): 11-16.
Patel, A. D. and Makawana, M. G. 2002. Genetic association in rustica tobacco (Nicotiana rustica L.). Tob. Res. 32(2): 45-49.
PTB (Pakistan Tobacco Board). 2016. Statistics, http://ptb.gov.pk/?q=node/33, accessed September 19th, 2016.
Shoukat, A.M., A.S. Mahboob, A.A Bashir and M. Afzal. 2007. Study of genetic parameters in segregating populations of spring wheat. Pak. J. Bot. 39(7): 2407-2413.
Shukla, S., A. Bhargava, A. Chatterjee and S.P Singh. 2004. Estimates of genetic parameters to determine variability for foliage yield and its different quantitative and qualitative traits in vegetable amaranth (A.tricolor). J. Genet. Breed. 58: 169-176.
Singh, R.K. and B.D. Chaudhary. 1985. Biometrical methods in quantitative genetic analysis. Kalyani Publishers, New Delhi, India.
Steel, R.G.D., J.H. Torrie and D.A. Discky, 1996. Principles and Procedures of Statistics: A Biometrical Approach, 3rd edition. McGraw Hill Book Co., New York, USA.
Taj, F.H. 1994. Miscellaneous crops. In: Bashir, E and B. Robyn (eds) Crop Production. National Book Foundation, Islamabad, Pakistan.
Zhang, H.Y., X.Z. Liu, C. Sheng and Y.M. Yang. 2008. Genetic Diversity among flue-cured tobacco cultivars based on RAPD and AFLP markers. Braz. Arch. Biol. Technol. 51(6): 1097-1101. https://doi.org/10.1590/S1516-89132008000600003
To share on other social networks, click on any share button. What are these?


